 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Нуклеозиды, нуклеотиды и нуклеиновые кислоты, их структура и свойства
|
|
Нуклеозиды и нуклеотиды являются продуктами гидролиза нуклеиновых кислот, но они присутствуют в живых организмах также в несвязанном состоянии, выполняя исключительно важную роль в обмене веществ.
Нуклеозиды - это природные гликозиды гетероциклических азотистых оснований (пиримидиновых и пуриновых), которые связаны с пентозами через атом азота. В зависимости от природы углеводного остатка (пентозы) различают рибонуклеозиды и дезоксирибонуклеозиды.

Названия нуклеозидов производятся от тривиального названия соответствующего гетероциклического азотистого основания с суффиксами -идин у пиримидиновых и -озин у пуриновых нуклеозидов. Исключение их этого правила сделано для нуклеозидов тимина.
| Компоненты нуклеозида | Нуклеозид | Индекс нуклеоаида |
| Аденин + рибоза | Аденозин | А |
| Аденин + дезоксирибоза | Дезоксиаденозин | dA |
| Гуанин + рибоза | Гуанозин | Г |
| Гуанин + дезоксирибоза | Дезоксигуанозин | dГ |
| Цитозин + рибоза | Цитидин | С |
| Цитозин + дезоксирибоза | Дезоксицитидин | dC |
| Урацил + рибоза | Уридин | u |
| Урацил + дезоксирибоза | Дезоксиуридин | dU |
| Тимин + рибоза | Риботимидин | Т |
| Тимин + дезоксирибоза | Тимидин | dT |
Нуклеозиды сокращенно чаще обозначают однобуквенным индексом, но существует также система трехбуквенного индекса.
Нуклеозиды, являясь N-гликозидами, устойчивы к гидролизу в слабощелочной среде, но расщепляются в кислой среде. Пуриновые нуклеозиды гидролизуются легко, пиримидиновые — труднее.
 |
Нуклеотиды являются сложными эфирами нуклеозидов и фосфорной кислоты, которая обычно этерифицирует гидроксогруппы при С-5' пентозы. В связи с наличием в молекуле остатка фосфорной кислоты нуклеотиды проявляют свойства двухосновной кислоты с p Ка 1 = 0,9 - 1,5 и p Ка 2 = 6 - 6,5.
Нуклеотиды называют или как соответствующие кислоты (монозамещенные производные фосфорной кислоты), или как соли (монофосфаты) с указанием в обоих случаях положения фосфатного остатка:

Поскольку с учетом значений  и
и  нуклеотиды в физиологических условиях находятся в ионизованном состоянии, последний способ предпочтительнее.
нуклеотиды в физиологических условиях находятся в ионизованном состоянии, последний способ предпочтительнее.
Большое значение в живых системах играют нуклеотиды, содержащие в своем составе ди- и трифосфатные группировки. Важнейшими среди этих производных являются аденозинди-фосфат (АДФ) и аденозинтрифосфат (АТФ), которые способны к взаимопревращениям путем наращивания или отщепления фосфатных групп:
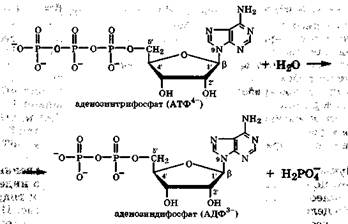 |
В этих соединениях фосфатные группы в физиологических условиях почти полностью ионизованы, поэтому их часто записывают в виде анионов АТФ4-, АДФ3-. Главная особенность этих нуклеотидов состоит в том, что их полифосфатные группы содержат одну или две ангидридные группы  . При гидролизе такой группы разрывается связь, называемая макроэргической, и выделяется около 33 кДж/моль (разд. 4.5). Именно с этим связана роль АТФ в клетке как поставщика химической энергии для биохимических и физиологических процессов.
. При гидролизе такой группы разрывается связь, называемая макроэргической, и выделяется около 33 кДж/моль (разд. 4.5). Именно с этим связана роль АТФ в клетке как поставщика химической энергии для биохимических и физиологических процессов.
При участии АТФ в организме также осуществляется реакция фосфорилирования гидроксилсодержащих соединений с образованием сложных эфиров фосфорной кислоты:

При фосфорилировании карбоксилсодержащих соединений образуются ацилфосфаты, которые содержат ангидридную группировку 

Фосфорилированные производные выступают активными метаболитами во многих биохимических процессах.
Нуклеотидами являются многие коферменты, например НАД-и ФАД-производные аденозинфосфата, а коэнзим А - производное аденозиндифосфата. Важнейшая биологическая роль нуклеотидов заключается в том, что из них построены нуклеиновые кислоты (полинуклеотиды).
Нуклеиновые кислоты в живых организмах играют главную роль в передаче наследственных признаков (генетической информации) и управлении процессом биосинтеза белка. Нуклеиновые кислоты - высокомолекулярные соединения с молекулярной массой от 20 тысяч до десятка миллиардов. Их полимерные цепи построены из мономерных звеньев-нуклеотидов. Особенность нуклеотидного звена заключается в том, что оно представляет собой трехкомпонентное образование, состоящее из гетероциклического азотсодержащего основания, углеводного компонента - пентозы - и фосфатной группы. Каркас полимерной цепи состоит из чередующихся пентозных и фосфатных остатков, связанных сложноэфирными связями (у С-3' и С-5'), а гетероциклические основания являются «боковыми» группами, присоединенными к пентозным остаткам за счет N-гликозидной связи:

Первичная структура нуклеиновых кислот определяется природой и последовательностью нуклеотидных звеньев, связанных сложноэфирными связями между пентозами и фосфатными группами (рис. 23.2).
Вторичная структура нуклеиновых кислот. Согласно вторичной структуре полинуклеотидная цепь представляет собой двойную спираль, в которой пуриновые и пиримидиновые основания направлены внутрь. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи имеются водородные связи, стабилизирующие такую структуру. Основания, образующие пары, связанные водородными связями, называются комплементарными. В ДНК комплементарными будут: аденин - тимин, образующие между собой две водородные связи, и гуанин - цитозин, связанные тремя водородными связями (рис. 23.3). Это означает, что пуриновым основаниям аде-нину и гуанину в одной цепи будут соответствовать пиримидиновые основания тимин и цитозин в другой цепи. Полинуклеотидные цепи, образующие двойную спираль, не идентичны, но комплементарны между собой.

Рис. 23.3. Комплементарные пары гетероциклических оснований и двойная спираль ДНК
Комплементарность цепей и последовательность звеньев составляют химическую основу важнейших функций нуклеиновых кислот: ДНК - хранение и передача наследственной информации, а РНК - непосредственное участие в биосинтезе белка. Молекулярная масса ДНК варьирует от нескольких миллионов до десятка миллиардов, у РНК - от десятка тысяч до нескольких миллионов.
Молекула ДНК, в отличие от молекулы РНК, в большинстве случаев состоит из двух комплементарных взаимозакрученных цепей. В зависимости от длины витка и угла спирали, а также ряда других ее геометрических параметров, различают более десяти разнообразных упорядоченных спиральных структур ДНК. В стабилизации этих структур наряду с водородными связями, действующими поперек спирали, большую роль играют межмолекулярные взаимодействия, направленные вдоль спирали между соседними пространственно сближенными азотистыми основаниями. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, их называют стэкинг-взаимодействиями. Таким образом, взаимодействия азотистых оснований между собой скрепляют двойную спираль молекулы ДНК и вдоль, и поперек ее оси.
Сильное стэкинг-взаимодействие всегда усиливает водородные связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, полярные группы которого находятся на поверхности спирали. При ослаблении стэкинг-взаимодействия молекулы воды, проникая внутрь спирали, конкурентно взаимодействуют с полярными группами оснований, инициируют дестабилизацию и способствуют дальнейшему распаду двойной спирали. Все это свидетельствует о динамичности вторичной структуры ДНК под воздействием компонентов окружающего раствора.
 |
Биспиральные структуры в молекулах РНК возникают в пределах одной и той же цепи в тех зонах, где расположены комплементарные азотистые основания аденин - урацил и гуанин -цитозин (рис. 23.4). В результате вторичная структура молекулы РНК содержит биспиральные участки и петли, число и размеры которых определяются первичной структурой молекулы и составом окружающего раствора.
Рис. 23.4. Вторичная структура молекулы РНК
Третичная структура нуклеиновых кислот. Двойная спираль молекул ДНК существует в виде линейной, кольцевой, суперкольцевой и компактных клубковых форм. Между этими формами совершаются взаимные переходы при действии особой группы ферментов - топоизомераз, изменяющих пространственную структуру (рис. 23.5).
Третичная структура многих молекул РНК пока еще требует окончательного выяснения, но уже установлено, что она зависит не только от первичной и вторичной структуры, но и от состава окружающего раствора.
Биологические функции и ДНК, и РНК полностью определяются только совокупностью первичной, вторичной и третичной структур. При этом следует отметить, что стабилизация вторичной и третичной структур нуклеиновых кислот, так же как у белков, происходит за счет ассоциации по принципу самоорганизации под влиянием и при участии компонентов окружающего раствора, и прежде всего молекул воды.

Рис. 23.5. Третичная структура молекулы ДНК: а - линейная, б -кольцевая, в -суперкольцевая, г -компактный клубок
Поверхностные свойства. Макромолекулы нуклеиновых кислот состоят из полярных групп, и поэтому их поверхность достаточно гидрофильна. Вследствие этого в водных растворах нуклеиновые кислоты при их малой концентрации, низкой молекулярной массе и при достаточно большой концентрации свободных молекул воды самопроизвольно образуют истинные растворы, а в случае большой молекулярной массы - лиофильные коллоидные растворы.
Наличие на поверхности макромолекул нуклеиновых кислот отрицательного заряда, возникающего за счет диссоциации фосфатных групп, способствует образованию ассоциативных комплексов - нуклеопротеинов, состоящих из нуклеиновых кислот и основных белков (рI > 8,0).
Учет только полярности заместителей в молекуле нуклеиновых кислот при описании их поверхностных свойств явно недостаточен, так как состав и последовательность азотистых оснований их макромолекул несут наследственную информацию живого организма. При синтезе дочерних нуклеиновых кислот на исходных полинуклеотидах и при синтезе белка поверхность полинук-леотидов используется как информационная матрица. Именно эта особенность нуклеиновых кислот определяет их уникальную биологическую роль в обеспечении жизненных процессов.
Структурно-информационные свойства. Нуклеиновые кислоты - информационные биополимеры, осуществляющие хранение и передачу генетической информации во всех живых организмах, а также участвующие в биосинтезе белков. ДНК является носителем генетической информации, которая записана через определенную последовательность расположения в цепи четырех гетероциклических оснований. Первый этап реализации генетической информации заключается в том, что на конкретных участках одной из нитей молекулы ДНК происходит синтез молекул РНК. Биосинтез РНК, называемый транскрипцией, обычно происходит в результате комплементарного копирования ДНК-матрицы с помощью РНК-полимеразы. Синтезированная РНК содержит точную копию конкретного участка ДНК.
В результате транскрипции образуются четыре различных вида РНК: рибосомалъная рРНК, матричная мРНК (информационная), транспортная тРНК и малые ядерные РНК, роль которых разнообразна, но до конца еще не выяснена. Каждая из синтезированных РНК играет строго определенную роль на втором этапе реализации генетической информации - трансляции. Реализация генетической информации с помощью нуклеиновых кислот происходит по схеме:

Рибосомальная РНК входит совместно с белками в состав рибосом. Матричная РНК, объединяясь с рибосомами, образует полирибосому, в которой с помощью ферментов и транспортных РНК, поставляющих определенные аминокислоты, происходит трансляция - синтез белков в соответствии с информацией, записанной на мРНК. Информация о последовательности аминокислот в молекуле белка считывается с последовательности гетероциклических оснований в мРНК. Конкретная группа из трех гетероциклических оснований в молекуле нуклеиновой кислоты, которая соответствует отдельной аминокислоте, называется кодоном. Совокупность кодонов составляет генетический код. Генетический код един для всего живого: у любого вида организмов каждая из ос-аминокислот кодируется одним и тем же кодоном или одними и теми же кодонами. (Несколько кодонов могут кодировать одну и ту же аминокислоту, но один и тот же кодон не способен кодировать разные аминокислоты.)
Жидкокристаллическое состояние нуклеиновых кислот. Рассмотрев структуру нуклеиновых кислот, убеждаешься в том, что для них характерна определенная ориентационно-пространственная организация нуклеотидов. Каждый нуклеотид анизотропен, а с образованием комплементарной пары в цепи анизотропные свойства системы усиливаются, следовательно, при рассмотрении двойной спирали всей молекулы ДНК роль анизотропии становится очень существенным фактором для описания ее свойств. Поэтому для молекул ДНК, молекулярная масса которых достигает 109, вполне реально, что в растворе отдельные достаточно крупные фрагменты этой строго организованной гигантской молекулы могут находиться в жидкокристаллическом состоянии, образуя внутримолекулярные термотропные жидкие кристаллы (при определенной температуре) или лио-тропные жидкие кристаллы (при определенной степени гидратации рассматриваемого фрагмента). Число таких фрагментов и их ориентация в пространстве сильно влияют на состояние ДНК в клетке и ее биологические функции.
Кроме того, различные лиотропные жидкокристаллические состояния могут самопроизвольно формироваться в системах полинуклеотиды - вода или нуклеопротеиды - вода в соответствии со свойствами лиофильных коллоидных растворов. В таких растворах могут происходить множественные переходы из одного жидкокристаллического состояния в другое, которые изменяют биологические функции соответствующих систем и совершаются под действием направленного поля или самопроизвольно. В настоящее время установлено, что внутри- и межмолекулярные жидкокристаллические образования нуклеиновых кислот или их комплексов с белками играют важную роль в процессах передачи информации и биосинтеза новых нуклеиновых кислот и белков на молекулярном уровне.
Денатурация. Подобно денатурации белков происходит денатурация нуклеиновых кислот, сопровождаемая разрушением их третичной и вторичной структур и сохранением первичной структуры. Это происходит под влиянием тех же факторов, что и в случае белков, но интенсивность фактора в случае нуклеиновых кислот, естественно, может быть другой, чем при денатурации белка. Под воздействием того или иного фактора снижается прочность водородных связей и уменьшается эффективность стэкинг-взаимодействия между азотистыми основаниями в макромолекуле. Это способствует раскручиванию двухцепочечных спиралей с образованием неупорядоченных одноцепочечных клубков. Поскольку при денатурации сохраняется первичная структура нуклеиновых кислот, то данный процесс может иметь обратимый характер.
Процесс денатурации нуклеиновых кислот разделяют на две стадии. На первой стадии две цепи частично раскручиваются, но остаются соединенными хотя бы в одном небольшом участке. На второй стадии две цепи полностью отделяются друг от друга. Первая стадия легко обратима. После второй стадии ре-натурация протекает очень медленно, особенно в случае ДНК с большой молекулярной массой.
Кислотно-основные свойства. Сильнополярные фосфатные группы нуклеиновых кислот характеризуются значением  Таким образом, нуклеиновые кислоты - это довольно сильные поликислоты, полностью ионизованные при рН выше 4, и поэтому их поверхность несет отрицательный заряд. Именно это обстоятельство объясняет большую склонность нуклеиновых кислот к взаимодействию с полиаминами, у которых между атомами азота содержатся две или три метиленовые группы. Однако особый интерес вызывает кислотно-основное взаимодействие нуклеиновых кислот с белками, которые являются полиамфолита-ми, образуя комплексные ассоциаты (соли), называемые нуклеопротеинами. Особенно активно нуклеиновые кислоты взаимодействуют с основными белками (рI > 8), имеющими в нейтральной среде в основном положительный заряд. Так, ДНК образует прочный комплекс с белками-гистонами, входящими в состав хромосом. Гистоны содержат 25-30 % остатков лизина и аргинина, основные функциональные группы которых при рН = 7 заряжены положительно. Они, электростатически взаимодействуя с отрицательно заряженными фосфатными группами, расположенными на периферии двойной спирали ДНК, образуют достаточно прочный комплексный ассоциат, в котором структура ДНК дополнительно стабилизирована. При ослаблении связей между ДНК и гистоном в силу тех или иных причин, например в результате изменения ионной силы среды, происходит дестабилизация ДНК. Этим и определяется регуляторная роль гистонов в функционировании генома.
Таким образом, нуклеиновые кислоты - это довольно сильные поликислоты, полностью ионизованные при рН выше 4, и поэтому их поверхность несет отрицательный заряд. Именно это обстоятельство объясняет большую склонность нуклеиновых кислот к взаимодействию с полиаминами, у которых между атомами азота содержатся две или три метиленовые группы. Однако особый интерес вызывает кислотно-основное взаимодействие нуклеиновых кислот с белками, которые являются полиамфолита-ми, образуя комплексные ассоциаты (соли), называемые нуклеопротеинами. Особенно активно нуклеиновые кислоты взаимодействуют с основными белками (рI > 8), имеющими в нейтральной среде в основном положительный заряд. Так, ДНК образует прочный комплекс с белками-гистонами, входящими в состав хромосом. Гистоны содержат 25-30 % остатков лизина и аргинина, основные функциональные группы которых при рН = 7 заряжены положительно. Они, электростатически взаимодействуя с отрицательно заряженными фосфатными группами, расположенными на периферии двойной спирали ДНК, образуют достаточно прочный комплексный ассоциат, в котором структура ДНК дополнительно стабилизирована. При ослаблении связей между ДНК и гистоном в силу тех или иных причин, например в результате изменения ионной силы среды, происходит дестабилизация ДНК. Этим и определяется регуляторная роль гистонов в функционировании генома.
Рибонуклеиновые кислоты также образуют с белками нуклеопротеины. Так, рибосомы состоят из 50-65 % рибосомной РНК и 35-50 % белков, содержащих до 25 % основных аминокислот. Масса одной рибосомной субъединицы составляет несколько миллионов, а диаметр 1,8 • 10-6 м. При контакте с 0,5-1,0 М растворами солей при низкой температуре происходит отделение белка от РНК в рибосомах вследствие их дегидратации. Аналогичная диссоциация происходит при увеличении рН до 12 из-за изменения заряда белковой молекулы.
Вирусы представляют собой устойчивые комплексные ассо-циаты, содержащие до 30 % нуклеиновой кислоты и большое число белковых молекул, уложенных в определенном порядке и образующих специфическую трехмерную структуру. В состав вируса может входить как ДНК, так и РНК.
Кислотно-основные свойства нуклеиновых кислот обусловлены не только наличием фосфатных групп, но и присутствием азотистых оснований. Азотистые основания нуклеиновых кислот, как было показано в разд. 23.3, являются амфолитами. Вследствие того, что у них и основные и кислотные свойства выражены слабо, внутримолекулярного солеобразования, как у аминокислот, в нуклеиновых кислотах не происходит. Кислотно-основные свойства гетероциклических оснований влияют главным образом на состояние и прочность водородных связей и стэкинг-взаимодействий, возникающих между ними. Поскольку на эти виды взаимодействий сильно влияет рН среды, изменение конформации нуклеиновых кислот может происходить при незначительном изменении рН.
Окислительно-восстановительные свойства. Нуклеиновые кислоты не содержат групп, склонных к окислительно-восстановительным превращениям при мягком воздействии. Поэтому они относительно устойчивы к воздействию мягких окислителей и восстановителей. При жестком окислении в водной среде нуклеиновые кислоты превращаются, как все органические соединения в организме, в СО2 и Н2О, а из-за присутствия в их составе атомов азота образуют мочевую кислоту, мочевину или соли аммония; кроме того, из-за наличия фосфатных групп образуются неорганические фосфаты.
Комплексообразующие свойства. Нуклеиновые кислоты являются активными полидентатными лигандами, содержащими как "жесткие" центры - ионизованные фосфатные группы, так и "мягкие" центры - полярные группы 
азотистых оснований. За счет "жестких" фосфатных центров нуклеиновые кислоты образуют малоустойчивые комплексы с очень "жестким" катионом К+ и более прочные комплексы с катионами Mg2+ и Са2+. "Мягкие" центры расположены на гетероциклических основаниях, и за счет их образуются прочные комплексы с "мягкими" катионами d-металлов. Образование комплексных соединений нуклеиновых кислот с катионами металлов, естественно, приводит к изменению их конформации, а следовательно, и их химической и биологической активности.
Глава 24
Дата публикования: 2014-10-16; Прочитано: 9117 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
