 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Биосинтез белков (трансляция)
|
|
Биосинтез белков в организмах осуществляется при помощи рибосом и является матричным синтезом наряду с репликацией и транскрипцией и носит название трансляции. Матрицей в процессе трансляции служит мРНК. Трансляция отличается от репликации и транскрипции двумя особенностями:
- нет соответствия между числом мономеров в матрице (мРНК) и в продукте реакции (белке);
- матрица (мРНК) и образующаяся на ней полипептидная цепь (белок) не комплементарны друг к другу.
Это означает, что механизм использования матрицы при трансляции иной, чем при репликации и транскрипции.
Первичная структура мРНК содержит в себе биологический код (информацию о первичной структуре белков), и в процессе трансляции происходит как бы декодирование этой информации и образование соответствующего белка.
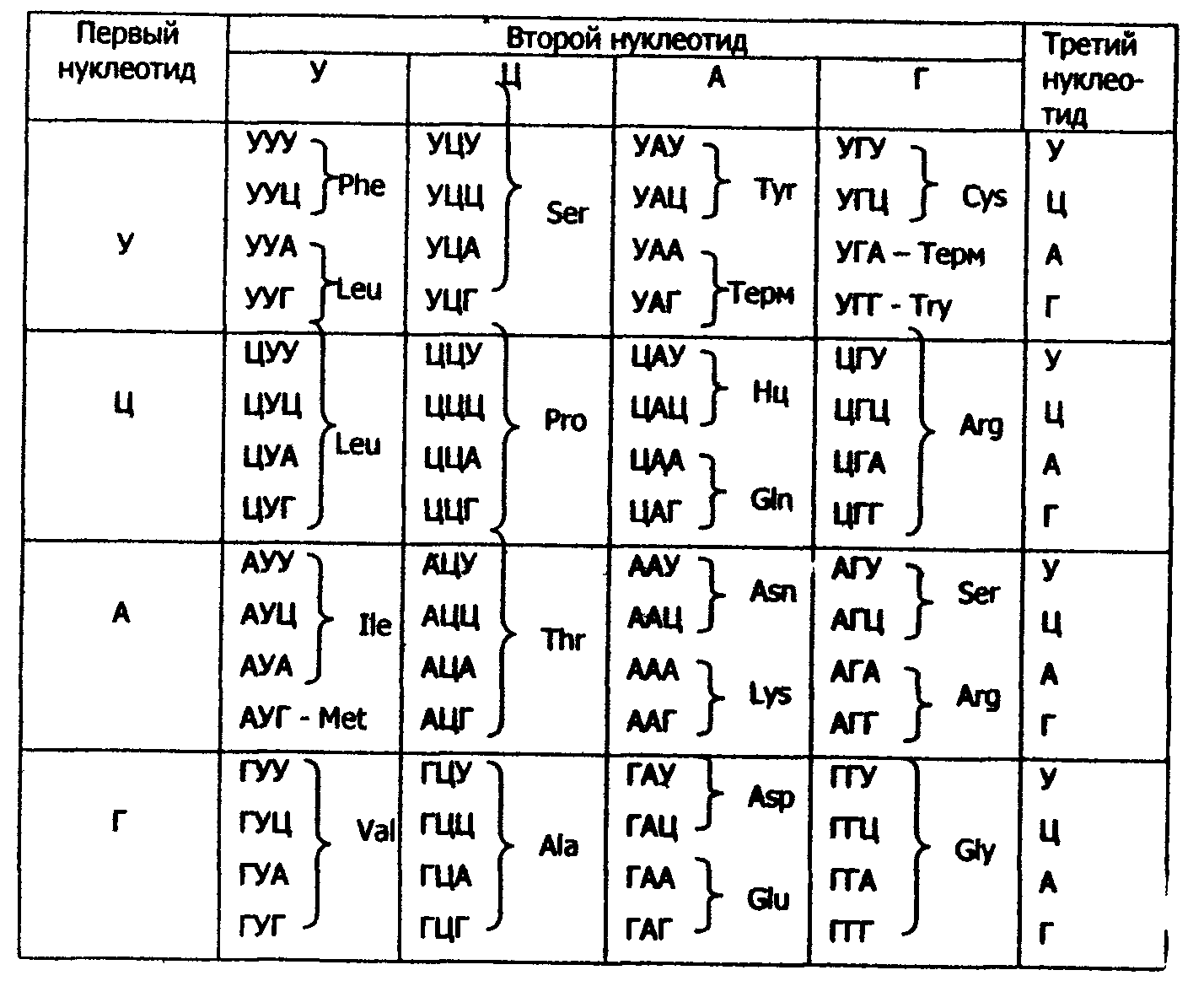
Установлено, что в биологическом коде кодовое число равно трем, т.е. три нуклеотидных остатка (триплет, или кодон) кодируют включение в белок одной аминокислоты. Число разных триплетов равно 43=64 (4 - число нуклеотидных мономеров в РНК). 61 триплет используется для кодирования аминокислот, а три: УАА, УАГ, УГА - являются терминирующими - на них обрывается дальнейшее наращивание пептидной цепи белка (табл.15).
Каждый триплет кодирует только одну аминокислоту (свойство специфичности кода); с другой стороны, одна аминокислота может кодироваться двумя и более (до 6) триплетами (свойство вырожденности кода). Например, включение в белок серина кодируется шестью триплетами: УЦУ, УЦЦ, УЦА, УЦГ, АГУ и АГЦ (см. табл. 15). Установлено, что биологический код одинаков для всех живых организмов разной степени организации (свойство универсальности кода).
Путь информации от ДНК к белку представляется следующим образом:
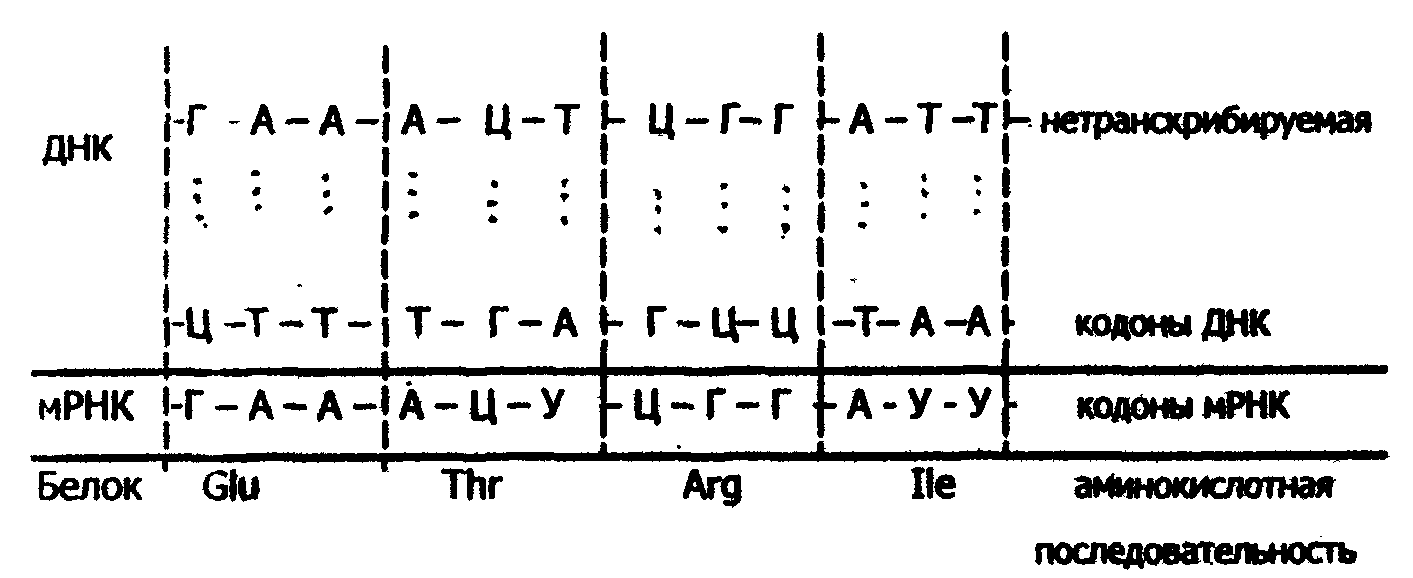
Таблица 15. Биологический код
Прямое комплементарное взаимодействие между аминокислотами и триплетами нуклеотидов в мРНК, как уже было сказано выше, невозможно. Взаимодействие косвенное. Роль посредника (или адаптера) в этом процессе выполняют молекулы тРНК: для каждой аминокислоты своя, что обозначается следующим образом: тРНКАла (аланиновая тРНК); тРНКВал (валиновая тРНК) и т.д. Поскольку код вырожденный, то число разных тРНК должно быть не менее 61.
Взаимодействие тРНК с аминокислотами - ферментативный процесс, идущий с поглощением энергии и приводящий к образованию ковалентной связи между аминокислотой и тРНК:
аминокистота+тРНКаминокислоты + АТФ → аа-тРНКаминокислоты +АМФ +Н4Р2О7.
Образуется соединение называемое аминоацил-тРНК (аа-тРНК). Аминокислота присоединяется к З'-концу нуклеотидной цепи тРНК (где имеется последовательность А-Ц-Ц, общая для всех тРНК). При этом получается сложноэфирная связь за счет карбоксильной группы аминокислоты, и гидроксильной группы концевого остатка адениловой кислоты в тРНК:
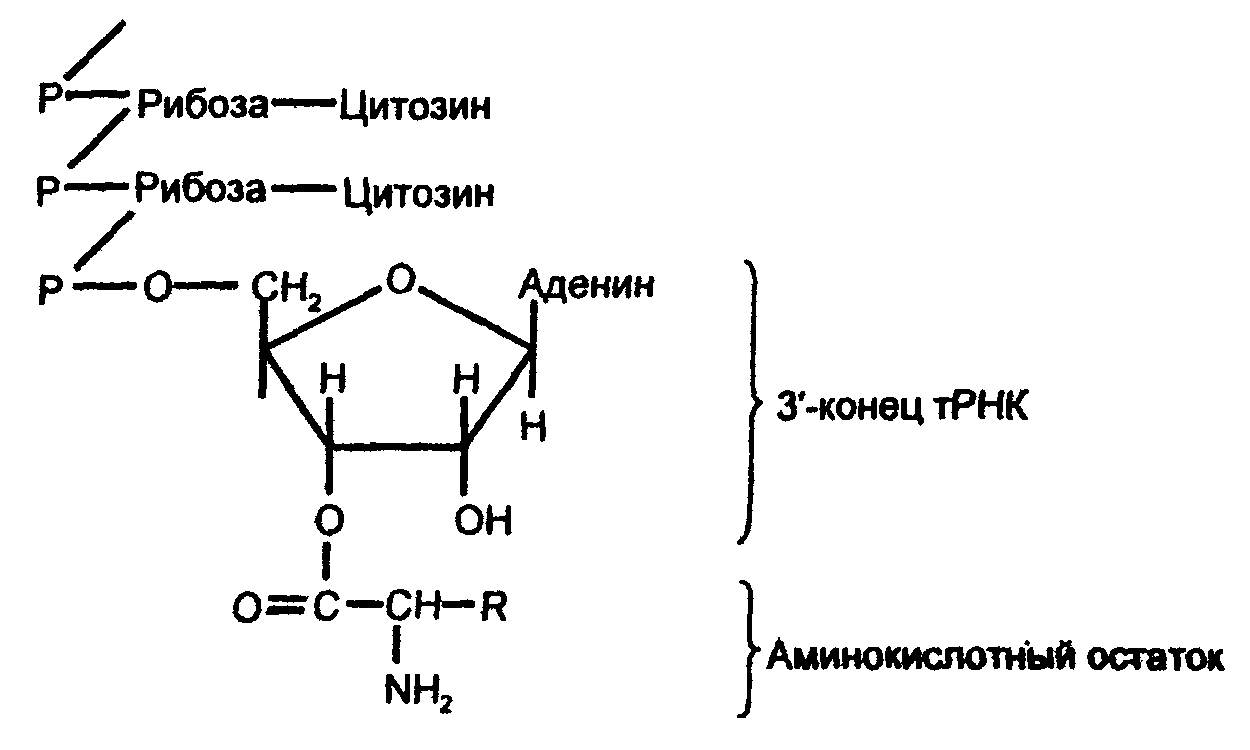
Эта связь является макроэргической, поэтому образование аа-тРНК можно рассматривать как активацию аминокислоты. Данная реакция катализируется ферментами аминоацил-тРНК-синтетазами, обладающими субстратной специфичностью: каждый фермент катализирует реакцию только одной аминокислоты с соответствующей ей тРНК.
Образовавшиеся аа-тРНК взаимодействуют с мРНК за счет комплементарности соответствующему кодону мРНК триплета нуклеотидов в одной из петель клеверного листа тРНК, называемого антикодоном.
Таким образом, аа-тРНК присоединяется антикодонами к соответствующим кодонам мРНК и образуется последовательность аминокислот в соответствии с расположением кодонов в мРНК - возникает белок с определенной первичной структурой.
Процесс активации аминокислот происходит в гиалоплазме клетки, а непосредственно синтез белка - на рибосомах.
Таким образом, для биосинтеза белка требуются: мРНК как генетическая матрица, содержащая информацию о первичной структуре белка; аминоацил-тРНК - для чтения «текста» мРНК и как источник аминокислот при сборке белка; рибосомы как молекулярные машины для последовательного соединения аминокислот в полипептидную цепь в соответствии с программой мРНК. Кроме того, используется ГТФ как источник энергии при синтезе белка в рибосомах; белковые «факторы», помогающие на разных фазах сборки белка в рибосомах, а также некоторые ионы как кофакторы ферментов (Мg2+, К+ и др.).
Устройство рибосом прокариотов и эукариотов примерно одинаково. Отличаются они лишь молекулярной массой. У эукариотов она больше, чем у прокариотов. Состоят рибосомы из двух субчастиц - большой и малой; скелет каждой из них образует рРНК, окруженная белками.
Установлено, что рибосомы активны только в полностью собранном виде. Рибосомы, не участвующие в синтезе белка, легко диссоциируют на субчастицы. Рибосомы могут свободно перемещаться в различные участки клетки, что дает возможность собирать белки в клетке там, где это нужно.
Весь процесс трансляции, протекающий на рибосомах, делят на три стадии: инициация (начало), элонгация (удлинение полипелтидной цеgи) и терминация (окончание).
Инициация. Начало трансляции - наиболее медленный процесс. В нерабочем состоянии субчастицы рибосом разомкнуты. мРНК, поступившая из ядра в цитоплазму, связывается с малой субчастицой на поверхности, обращенной к большой субчастице. Причем присоединение идет 5'-концом РНК, так как "чтение" программы РНК всегда идет в направлении 5' → 3'. В пределах субчастицы умещаются только два кодона мРНК. Первым кодоном мРНК у 5'-конца является АУГ(Меt) или ГУГ(VаI). Эти кодоны называют инициирующими, так как именно с них всегда начинается трансляция в рибосомах. Этим кодонам соответствует антикодон метионил-тРНКMet. Кроме того, в инициации участвуют внерибосомные белки (факторы инициации), которые облегчают связывание мРНК с малой субчастицой и ГТФ. К этому первичному комплексу (факторы инициации - малая субчастица- мРНК - Met -т PHKMet-ГТФ) присоединяется большая субчастица рибосом, после чего факторы инициации удаляются из рибосом. Необходимая для смыкания субчастиц энергия получается за счет гидролиза ГТФ. Образовавшийся инициирующий комплекс (мРНК -Met - TPHKMet- рибосома) готов к элонгации. Причем метионил - tPHKMet своим антикодоном комплементарно спаривается с кодоном АУГ мРНК, т.е. как бы "подвешивается" на водородных связях к мРНК, а акцепторный конец, где находится аминокислота, прикрепляется к большой субчастице рибосом (ее пептидильному центру).
Элонгация. Синтез полипептида всегда начинается от N-конца и заканчивается С-концом. Наращивание полипептида на одну аминокислоту осуществляется в три этапа.
1. Связывание аа-тРНК1. К инициирующему комплексу присоединяется аа-тРНК1, соответствующая первому кодону мРНК (следующему за инициирующим кодоном). Эта аа-тРНК1 взаимодействует и с мРНК (своим а тикодоном), и с определенными участками рибосомы - назовем их центр связывания. Этот процесс требует энергию одной молекулы ГТФ, а также участия в ней рибосомного белка - фактора элонгации EF1.
2. Образование пептидной связи. Остаток метионина с Met-тРНК1 переносится на аминогруппу остатка аминокислоты в аа-тРНК1. При этом получается дипептидил- тРНК1, связанная с кодоном 1 и с центром связывания.
3. Транслокация - перемещение рибосомы относительно мРНК и дипептидил-тРНК1. В результате этого дипептидил-тРНК1 оказывается в области пептидильного центра рибосомы, но по-прежнему связана с первым кодоном мРНК. При этом тРНКMet освобождается из комплекса. Транслокация требует энергию двух молегул ГТФ и внерибосомный белок - фактор элонгации EF2.
Далее эти три этапа повторяются, происходит наращивание пептидной цепи с огромной скоростью: за 1 минуту синтезируется пептид, содержащий 40-60 аминокислот.
Терминация. Удлинение пептидной цепи продолжается до тех пор, пока на пути рибосомы не встретится один из терминирующих триплетов мРНК: УАА, УАГ или УГА. В области этих триплетов при участии внерибосомных белков - факторов терминации - происходит гидролитическое расщепление связи между пептидом и последней тРНК, освобождается готовый белок.
На включение в белок каждой аминокислоты расходуется энергия четырех макроэргических молекул: одной АТФ (на стадии синтеза аа-РНК) и трех ГТФ (на стадии элонгации).
Рибосома в ходе трансляции перемещается вдоль мРНК в направлении 5' 3' -конец. По мере освобождения 5'-конца к нему присоединяется новая рибосома и т.д. На молекуле мРНК может поместиться несколько рибосом (каждая соответствует примерно 30 кодонам) - возникает полирибосома. Чех длиннее пептидная цепь кодируемого белка, тем длиннее молекула РНК и тем больше число рибосом в полирибосоме.
В процессе трансляции по мере удлинения пептидной цепи формируются сразу вторичная и третичная структуры белка. Функционально активный белок образуется окончательно в ходе посттрансляционной достройки.
Регуляция биосинтеза белков. Концентрация многих белков в клетке непостоянна и изменяется в зависимости от условий, например, в зависимости от количества и состава пищи, в процессе онтогенеза, при введении некоторых лекарственных веществ. Это происходит в результате регуляции скоростей синтеза и распада белков. На рис.40 указаны процессы, от которых зависит концентрация белков в клетке и которые могут быть точками приложения регуляторных механизмов.
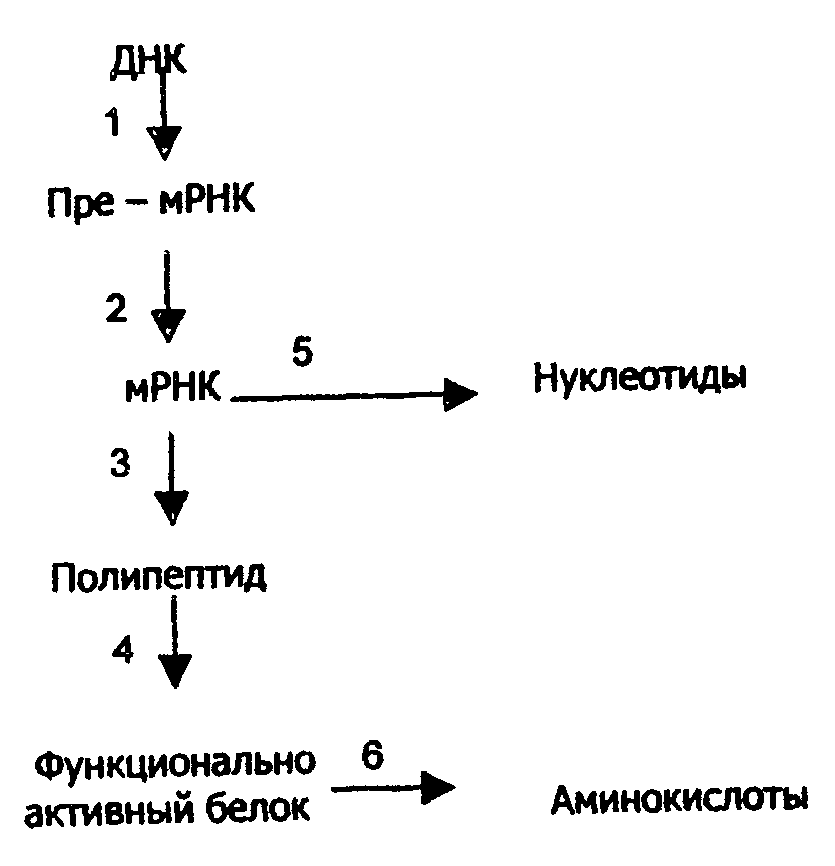
Рис. 40. Основные процессы, от скорости которых зависит концентрация белка в живой клетке: 1 транскрипция; 2 - созревание и транспорт мРНК из ядра в цитоплазму; 3 - трансляция; 4 - посттрансляционная достройка белка; 5 - распад мРНК; 6 - распад белка
Дата публикования: 2015-11-01; Прочитано: 2147 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
