 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
хвойные породы Л И Г Н И Н лиственные породы
|
|
 |  | ||||
 | |||||
|
|
|

|
Рис. 24. Структурные единицы лигнина
|
|
|
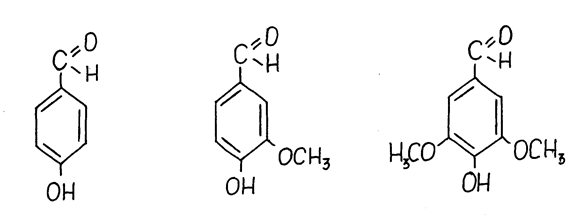
Рис. 25.Структурные формулы ароматических альдегидов
Природный лигнин в древесине – это пространственный сетчатый полимер. Основные типы связей между мономерными единицами: простые эфирные (С-О-С) и углеродные (С – С).
Гидролизуемую углеводную часть древесины условно подразделяют по скорости гидролитической деструкции в водных растворах минеральных кислот на легко – и трудногидролизуемые полисахариды.
В составе холоцеллюлозы к трудногидролизуемым полисахаридам относятся целлюлоза, как кристаллический полимер, и небольшая часть гемицеллюлоз, закристаллизованных совместно с целлюлозой и называемых целлюлозанами (рис. 26). К легкогидролизуемым полисахаридам относятся гемицеллюлозы, как аморфные полимеры, и небольшая часть целлюлозы (аморфная часть). В древесине в целом к легкогидролизуемым полисахаридам относятся также все водорастворимые нецеллюлозные полисахариды и полиурониды.

Рис.26. Схематическое изображение целлюлозанов в микрофибрилле целлюлозы
Экстрактивные вещества – это группа веществ, которые можно извлекать из древесины нейтральными растворителями (экстрагировать). Экстрактивные вещества подразделяют на несколько групп: вещества, летучие с паром (эфирные масла); вещества, растворимые в органических растворителях (смолы); вещества, растворимые в воде. Экстрактивные вещества, за исключением водорастворимых полисахаридов (крахмал, арабиногалактан) и полиуронидов (пектиновые вещества, камеди), представляют собой низкомолекулярные соединения. Содержание экстрактивных веществ зависит от породы древесины. Массовая доля экстрактивных веществ в некоторых породах древесины может составлять 3-4 %, они разнообразны по составу.
3.2. Химический состав древесины хвойных и лиственных пород
Древесина различных пород существенно различается по химическому составу. Химический состав древесины одной и той же породы не является строго постоянным и изменяется в зависимости от других факторов: условий и района произрастания, возраста дерева. Даже в стволе одного и того же дерева могут наблюдаться заметные различия в содержании отдельных компонентов по высоте ствола, по диаметру. Химический состав древесины хвойных и лиственных пород различен. Состав древесины наиболее важных отечественных пород приведен в табл. 2.
При анализе древесины данные анализа могут колебаться в довольно широких пределах, но в среднем древесина хвойных пород содержит больше лигнина (27-30 %) и меньше гемицеллюлоз (20-25 %), чем лиственная (соответственно, 18-24 % и 22-35 %). При этом хвойные породы содержат больше гексозанов и меньше пентозанов, чем лиственные. Содержание целлюлозы в древе-
Таблица 2
Химический состав древесины хвойных и лиственных пород (%)
| Древесные породы | Целлюлоза | Лигнин | Пентозаны | Гексозаны | Зола | Растворимые вещества | |
| В эфире | В воде | ||||||
| Сосна | 50,6 | 27,5 | 10,4 | 11,8 | 0,2 | 4,1 | 2,6 |
| Ель | 45,2 | 29,0 | 9,5 | 12,3 | 0,3 | 2,5 | 1,9 |
| Лиственница | 36,2 | 26,1 | 7,8 | 11,8 | 1,0 | 1,8 | 20,0 |
| Пихта | 48,5 | 29,9 | 5,3 | 10,9 | 0,7 | 0,9 | 2,2 |
| Береза | 45,1 | 25,2 | 22,1 | 6,0 | 0,4 | 2,7 | 2,9 |
| Осина | 41,8 | 21,8 | 16,3 | 3,0 | 0,3 | 0,8 | 2,8 |
сине хвойных и лиственных пород примерно в одинаковых пределах (массовая доля от 35-50 %).
4. Строение и состав клеточной оболочки
4.1. Слои клеточной оболочки
Клеточная стенка (оболочка клетки) анатомических элементов древесины, волокон технической целлюлозы и других волокнистых полуфабрикатов имеет сложное строение.
Если рассмотреть отдельно взятую клетку в разрезе (рис. 27), то можно видеть, что оболочка клетки состоит из нескольких слоев: первичной стенки (Р) и вторичной стенки (S). Вторичная оболочка состоит из трех слоев: наружного (S1), среднего (S2), внутреннего (S3).

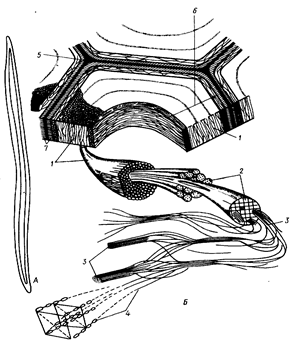
Рис. 27. Поперечный разрез трахеиды поздней древесины сосны. 4200-кратное увеличение: М - срединная пластинка; Р - первичная оболочка; S1, S2 и S3 - наружный, средний и внутренний слои вторичной оболочки
Клетки связываются между собой межклеточным веществом (М) или срединной пластинкой (истинной срединной пластинкой). В период развития клетки межклеточное вещество состоит из пектиновых веществ. В зрелой клетке после одревеснения клеточной стенки основным компонентом межклеточного вещества становится лигнин. Слой аморфен; его толщина составляет 0,2-1,0 мкм с утолщениями в углах клетки.
Первичная стенка (Р) – тонкий слой, являющийся единственной оболочкой, заключающей в себе протопласт (протоплазму и органоиды) в период увеличения поверхности клетки. Толщина слоя Р 0,1-0,3 мкм. Этот слой состоит из целлюлозы, гемицеллюлоз, пектиновых веществ, белков и лигнина, откладывающегося в период одревеснения. Лигнин появляется сначала в первичной стенке в углах клетки, а затем в межклеточном веществе и во всей первичной стенке.
Межклеточное вещество и первичные стенки двух смежных клеток тесно срастаются между собой, образуя сложную срединную пластинку (Р+М+Р).
В период утолщения клеточной стенки образуется вторичная стенка S за счет деятельности протоплазмы. Некоторые исследователи считают, что слой S3 следует рассматривать как индивидуальный слой – третичную стенку Т. Слои S1, S2, S3 (Т) различаются по толщине.
Слои S1 и S3 (Т) тонкие, а слой S2 толстый слой. Средний слой вторичной оболочки составляет большую часть клеточной стенки. Средний слой в трахеидах ранней древесины ели составляет 74-84 % от объема клеточной стенки (табл. 3).
Таблица 3
Толщина и соотношение слоев в оболочках трахеид ранней древесины ели
| Слой | Толщина, мкм | Содержание слоев, % |
| Р S1 S2 S3 | 0,23-0,34 0,12-0,35 1,77-3,68 0,10-0,15 | 7,0-14,2 5,2-10,8 73,3-84,0 2,7-4,2 |
У поздних трахеид толщина стенки (оболочки) увеличивается за счет S2, доля которого возрастает и соответственно уменьшается доля остальных слоев. Кроме перечисленных слоев в трахеидах древесины хвойных пород имеется тонкий (0,10-0,25 мкм) бородавчатый слой W, выстилающий полость клетки (люмен) L.
Строение клеточных стенок волокон либриформа древесины лиственных пород примерно такое же, как у трахеид поздней древесины и распределение слоев приблизительно следующее Р: S1: S2: S3 = 1: 10: 87: 2. Слой W встречается не у всех лиственных пород. По строению клеточных стенок паренхимные клетки и сосуды отличаются от трахеид и волокон либриформа.
4.2. Распределение компонентов древесины в клеточной оболочке
Основными структурными компонентами клетки являются целлюлоза, лигнин и гемицеллюлозы.
Молодые клеточные оболочки древесины состоят, главным образом, из целлюлозы и гемицеллюлоз. С течением времени, еще при жизни клетки, оболочки подвергаются одревеснению, так как в них накапливается лигнин, который делает клетки более устойчивыми против набухания, гидрофобными. Предполагают, что процесс накопления лигнина – результат жизнедеятельности протопласта клетки. Накопление лигнина является признаком старения клетки, и, возможно, причиной прекращения ее жизнедеятельности.
Изучение распределения компонентов древесины в клеточной стенке представляет очень трудную задачу. Результаты, полученные разными исследователями, несколько расходятся.
Большинство исследователей считают, что сложная срединная пластинка на 60-90 % состоит из лигнина (в ранней древесине примерно 70 %, а в поздней 80 %). Однако, слой тонкий и количество лигнина сложной срединной пластинки соответствует только 10-20 % общего количества лигнина всей клетки.
По направлению к полости клетки доля лигнина уменьшается.
 Слой S2 вторичной оболочки содержит наибольшее количество лигнина клетки. Его доля у хвойных пород составляет 30 % массы слоя, а у лиственных 20 %. В слое S3 (Т) доля лигнина не более 10-12 % массы слоя.
Слой S2 вторичной оболочки содержит наибольшее количество лигнина клетки. Его доля у хвойных пород составляет 30 % массы слоя, а у лиственных 20 %. В слое S3 (Т) доля лигнина не более 10-12 % массы слоя.
Основная масса целлюлозы находится в слое S2 (около 85 % от углеводной части этого слоя). Меньше всего целлюлозы в первичной стенке, примерно 40 % углеводной части, т.е. менее 12 % массы слоя. В слое S3 (Т) содержание целлюлозы невелико.
На рис. 28 приведена схема распределения основных компонентов клеточной оболочки.
 Распределение гемицеллюлоз и других нецеллюлозных полисахаридов изучено меньше, чем распределение целлюлозы и лигнина. Установлено, что в слоях S1 и S3 (Т) относительное содержание гемицеллюлоз выше, чем в слое S2, причем наибольшая концентрация гемицеллюлоз наблюдается в слое S1.
Распределение гемицеллюлоз и других нецеллюлозных полисахаридов изучено меньше, чем распределение целлюлозы и лигнина. Установлено, что в слоях S1 и S3 (Т) относительное содержание гемицеллюлоз выше, чем в слое S2, причем наибольшая концентрация гемицеллюлоз наблюдается в слое S1.
4.3. Строение целлюлозных микрофибрилл
В целлюлозе очень важное значение имеют водородные связи. В макромолекулах целлюлозы большое количество свободных гидроксильных групп (-ОН), которые способны образовывать водородные связи между звеньями и между макромолекулами целлюлозы (рис. 29). Водородные связи определяют физическую структуру целлюлозы (форму макромолекул, фазовые и физические состояния, надмолекулярную структуру) и оказывают влияние на все свойства целлюлозы – физические, физико-химические и химические (реакционную способность). 
 У целлюлозы в твердом состоянии возникает регулярная система водородных связей и, вследствие этого, кристаллическая решетка, образуются элементы надмолекулярной структуры – микрофибриллы, фибриллы, ламеллы и клеточная стенка в целом.
У целлюлозы в твердом состоянии возникает регулярная система водородных связей и, вследствие этого, кристаллическая решетка, образуются элементы надмолекулярной структуры – микрофибриллы, фибриллы, ламеллы и клеточная стенка в целом.
Основными элементами надмолекулярной структуры целлюлозы считают микрофибриллы, которые могут быть собраны в более крупные агрегаты - фибриллы (макрофибриллы) и распадаться на более тонкие элементы – элементарные фибриллы.
Существуют две основные концепции тонкой структуры микрофибрилл: двухфазной (аморфно-кристаллической системы) и однофазной кристаллической системы с дефектами кристаллической решетки. Существующие методы рентгеноструктурного анализа и расшифровки рентгенограмм не позволяют совершенно точно установить содержание малоупорядоченной части и отнести ее однозначно к аморфным участкам или дефектам кристаллической решетки.
Согласно теории аморфно-кристаллического строения целлюлозы, ее цепные макромолекулы проходят в продольном направлении микрофибрилл, образуя чередующиеся участки, различающиеся степенью упорядоченности, кристалличности, - кристаллические и аморфные в соответствии со схемой «бахромчатой фибриллы» (рис. 30).
Эти участки, если принять структурное понятие фазы, можно рассматривать как кристаллическую и аморфную фазы. Кристаллическая фаза постепенно переходит в аморфную. Каждая целлюлозная цепь проходит последовательно ряд кристаллических и аморфных участков, поскольку длина макромолекулы целлюлозы намного превышает длину кристаллических участков. Кристаллические участки также называют кристаллитами.

Рис. 30. Элементарные фибриллы с кристаллическими и аморфными участками: 1 – аморфный участок; 2 – кристаллический участок
В кристаллитах макромолекулы строго ориентированы, в них действует сильное межмолекулярное взаимодействие. В аморфных участках трехмерный порядок отсутствует и сохраняется лишь общая продольная направленность цепей.
Энергия межмолекулярного взаимодействия в аморфных участках меньше, чем в кристаллических, и поэтому аморфные участки более доступны для химических реагентов.
Целлюлоза относится к ориентированным полимерам, так как ее кристаллиты ориентируются в одном направлении, совпадающим с направлением микрофибрилл, которые в свою очередь, в слоях клеточной стенки идут по спиралям под определенным углом к оси волокна.
Взгляды различных исследователей на структуру микрофибрилл несколько различаются.
Согласно взглядам Фрей-Висслинга [6,7] микрофибриллы состоят из четырех элементарных микрофибрилл (рис. 31). Микрофибриллы имеют прямоугольное сечение размерами 10 х 20 нм. Они разделяются на четыре элементарные фибриллы (ультразвуком делигнифицированных целлюлозных волокон). Каждая элементарная фибрилла имеет в центре кристаллический стержень (3 х 7 нм), окруженный паракристаллической частью.

Рис. 31. Схема поперечного разреза микрофибриллы
4.4. Ультраструктура клеточной оболочки

 |
Оболочка одревесневшей растительной клетки состоит из нескольких слоев (Р, S1, S2, S3), которые отличаются друг от друга по своему происхождению, физическим свойствам и химическому составу, а также ориентацией и направлением целлюлозных микрофибрилл. Микрофибриллы в клеточной стенке имеют спиральное направление. Микрофибриллы, ориентированные в одном направлении, образуют тонкие слои ламеллы. На рис. 32 изображена схема расположения микрофибрилл целлюлозы в различных слоях клеточной оболочки.
Структура межклеточного вещества (М) аморфна, целлюлоза в нем практически отсутствует, однако предполагают, что одиночные микрофибриллы проходят через межклеточное вещество. В первичной стенке (Р) доля целлюлозы невелика и ее микрофибриллы образуют беспорядочную рыхлую сетку.
В слое S1 микрофибриллы образуют две группы перекрещивающихся спиралей, идущих почти перпендикулярно продольной оси волокна (от 700 до 900).
Концентрические слои в слое S2 состоят из микрофибрилл, ориентированных почти вдоль оси волокна (не более 300 к оси волокна). У различных видов растений угол различен.
Внутренний слой S3 очень тонкий. Считают, что в нем микрофибриллы образуют систему пересекающихся спиралей, идущих почти перпендикулярно оси волокна (700 – 800), но структура этого слоя более рыхлая, менее правильная, чем в слое S1, так как целлюлоза занимает в нем сравнительно небольшую часть.
На рис. 33 приведены микрофотографии расположения микрофибрилл в различных слоях клеточной стенки.

Рис. 33. Расположение микрофибрилл в слоях клеточной стенки
Целлюлозные микрофибриллы в клеточной стенке образуют каркас (фибриллярную арматуру), заключенный в лигноуглеводной (лигнингемицеллюлозной) матрице. Лигноуглеводная матрица представляет собой взаимное наложение трех сетчатых систем: сетчатой структуры, образованной химическими связями в лигнине; сетки, образованной химическим связями между лигнином и гемицеллюлозами; сетки, водородных связей в лигнине, в гемицеллюлозах и между ними.
В этой матрице лигнин аморфен и изотропен, а гемицеллюлозы аморфны, но у поверхностей целлюлозных микрофибрилл ориентированы в их направлении. Гемицеллюлозы частично могут находиться и внутри микрофибрилл между цепями целлюлозы. На рис.34 приведена схема последовательного распределения фибрилл, микрофибрилл, мицелл и участков молекул целлюлозы [8].
|
Таким образом, предполагаемая тонкая структура (ультраструктура) клеточной стенки (оболочки) допускает расщепление микрофибрилл при химической и механической обработке на продольные элементы различного размера.
Технология получения технической целлюлозы заключается в химической обработке (варке) древесинного вещества (древесины) варочными растворами при повышенной температуре и давлении. При варке происходит процесс делигнификации (удаление из древесинного вещества лигнина), который сопровождается частичным удалением гемицеллюлоз. В результате древесная ткань распадается на отдельные древесные волокна (клетки) с превращением последних в целлюлозные волокна.
При этом ультраструктура клеточной стенки существенно изменяется. Целлюлозные волокна становятся гибкими и эластичными. При последующем размоле целлюлозной массы происходит фибриллирование клеточных стенок – расщепление их на фибриллы и микрофибриллы, а при формовании бумаги фибриллярная структура позволяет связывать волокна между собой. Большую роль при этом играют водородные связи между макромолекулами целлюлозы.
5. Гетерокапиллярная структура древесины и особенности набухания целлюлозных волокон
Древесина имеет пористую гетерокапиллярную структуру. В ней существуют капиллярные пространства различных размеров.
Капиллярные пространства древесины подразделяют на две группы:
- капилляры первого порядка – микроскопические (межклеточные пространства, полости клеток и поры в стенках клеток);
- капилляры второго порядка – субмикроскопические (тонкие капилляры в клеточной стенке между ламеллами, фибриллами и внутри микрофибрилл).
Размеры капиллярных пространств меняются с возрастом дерева, содержанием влаги и т.д. Так, в насыщенной водой еловой древесине удельная площадь внутренней поверхности пространств первого порядка составляет около 0,2 м2/г, а пространств второго порядка – 300 м2/г.
Пористая структура древесины имеет большое значение для взаимодействия древесины с водой и передвижения воды и соков в дереве, а также для технологии переработки древесины – пропитки ее различными жидкостями, растворами химических реагентов, протекания процесса делигнификации при варке древесины.
Существование капиллярных пространств делает возможным набухание целлюлозных волокон. Особенности тонкого строения клеточных оболочек приводят к особенностям их набухания.
Освобожденные от лигнина целлюлозные волокна неодинаково набухают по длине и толщине. Так, в воде набухание в толщину достигает 20-30 %, а удлинение составляет лишь 1-2 %, а в растворе NaOH (17-18 %) набухание отдельного целлюлозного волокна по диаметру достигает 70 %, а в продольном направлении волокно даже укорачивается.
При набухании целлюлозных волокон в медноаммиачном реактиве, наблюдается образование характерных вздутий (бус), обусловленное особенностями ультраструктуры клеточной стенки (оболочки). Сначало волокно набухает равномерно, а затем в слоях Р и S1 образуются трещины, в которые проникает растворитель. Первичная стенка Р при этом отслаивается, спирально направленные микрофибриллы слоя S1 скользят по слою S2 и образуют перетяжки, а слой S2, набухая образует характерные вздутия (рис.35).
Рис. 35. Схема набухания целлюлозного волокна с образованием «бус»: 1 – перетяжка из микрофибрилл слоя S1; 2 - набухший слой S2; 3 - слой S3
Если слои P и S1 осторожно удалить, целлюлозное волокно будет набухать равномерно.
6. Пороки древесины [5].
Ложное ядро. При повреждении древесины в результате раневой реакции в ней образуется так называемое ложное ядро. Причиной данного явления могут быть грибы или бактерии, которые проникают в древесину через сломанные ветви и раны ствола. Ложное ядро может иметь разный цвет и форму. Оно образуется, например, в центральной части ствола, подобно настоящему ядру может быть в виде колец, а вокруг ядра – в виде небольших пятен или полос (рис. 36). Ложное ядро часто образуется у лиственных пород, у которых обычно центральная часть ствола бесцветная. Ложное ядро встречается у таких пород, как бук, береза, клен, осина, сосна.
 Раневая сердцевина (сердцевинные повторения). Раневая сердцевина напоминает настоящую сердцевину, но располагается в заболони и образуется вследствие ранения дерева. В месте травмы формируется раневая ткань, напоминающая сердцевину ствола (сосна).
Раневая сердцевина (сердцевинные повторения). Раневая сердцевина напоминает настоящую сердцевину, но располагается в заболони и образуется вследствие ранения дерева. В месте травмы формируется раневая ткань, напоминающая сердцевину ствола (сосна).
 В раневой сердцевине и в ложном ядре содержится большое количество экстрактивных веществ. В химической технологии переработки древесины (при варке целлюлозы) ложное ядро и раневая древесина вызывают трудности, как и обычная ядровая древесина.
В раневой сердцевине и в ложном ядре содержится большое количество экстрактивных веществ. В химической технологии переработки древесины (при варке целлюлозы) ложное ядро и раневая древесина вызывают трудности, как и обычная ядровая древесина.
Реактивная древесина. В древесине могут возникать изменения, обусловленные отклонением деревьев от вертикального положения по какой-либо причине (например, из-за сильных ветров, снега). Такую древесину называют реактивной. Реактивная древесина хвойных пород называется креневой. Она формируется в нижней части изогнутых стволов и ветвей. Реактивная древесина лиственных пород называется тяговой. Она образуется в  верхней части изогнутых стволов и вет
верхней части изогнутых стволов и вет
 |
вей (рис. 37).
Креневая древесина тяжелее, прочнее, плотнее и темнее обычной. В зоне креневой древесины образуются более широкие кольца прироста, между клетками образуются крупные межклетники (рис. 38).
|
 |
В креневой древесине по сравнению с нормальной содержится меньше целлюлозы и больше лигнина. У технической целлюлозы из-за присутствия креневой древесины снижаются прочностные свойства.
Тяговая древесина по сравнению с нормальной содержит меньше лигнина. Волокна этой древесины толстостенные, содержат желатинообразный слой. Тяговая древесина низкого качества. Выход целлюлозы за счет желатинообразного слоя выше, но ее прочностные свойства ниже.
Сучки. Ветви дерева отходят от центра ствола и поэтому часть их всегда размещается внутри дерева (рис. 39). У живой ветви эта часть, так называемый сучок, образует единую систему с живыми клетками ствола. Направление волокон сучка сильно отличается от направления волокон в стволе. Волокна ствола, окружающие сучок, также отличаются по направлению, они как бы закручены по спирали вдоль сучка. При варке древесины наличие сучков в древесине яв
|
 |
7. Микростроение и состав коры
Кора дерева состоит из двух основных частей: внутреннего слоя (луба) и наружного слоя (корки).
Ткани коры выполняют ряд функций: проводящую, запасающую и защитную. Ткани луба проводят соки вниз по стволу и хранят резервные питательные вещества, а ткани корки обеспечивают защиту от внешних воздействий.
Проводящая ткань луба состоит из ситовидных клеток у хвойных деревьев и ситовидных трубок у лиственных.
Ситовидные клетки – узкие длинные живые клетки, образующие продольные ряды и сообщающиеся между собой через пористые ситовидные поля в стенках клеток на их концах.
Ситовидные трубки образуются из клеток-члеников, сообщающиеся друг с другом через пористые (с более крупными порами) ситовидные пластинки на поперечных стенках.
Запасающую функцию выполняет лубяная паренхима, клетки которой образуют лубяные лучи, служащие продолжением сердцевинных лучей ксилемы. Присутствует также и вертикальная паренхима. Стенки ситовидных элементов и паренхимных клеток состоят преимущественно из целлюлозы и не лигнифицированы.
Механическую функцию выполняют склеренхимные клетки, к которым относят лубяные волокна и склереиды.
Лубяные волокна. Лубяные волокна похожи на волокна либриформа. У некоторых растений (лен, рами) лубяные волокна достигают большей длины.
Склереиды (каменистые клетки) – короткие клетки, стенки их сильно утолщены и одревеснели (лигнифицированы).
Наружный слой коры (корка) – состоит в основном из мертвых тканей.
Между лубом и коркой находится промежуточный слой паренхимных клеток (пробковый камбий). При делении клеток этого камбия в сторону луба откладываются клетки лубяной паренхимы, а в сторону корки -–пробковые клетки. Оболочки пробковых клеток не имеют пор и пропитаны особым жировым веществом суберином, делающим пробковую ткань малопроницаемой для воды и газов. С уберин – комплекс высокомолекулярных гидрокислот и фенольных кислот.
Пробковая ткань березы – береста.
По химическому составу кора деревьев значительно отличается от состава древесины. Внутренняя и наружная части коры, имеющие различное функциональное значение и соответственно строение, существенно отличаются друг от друга и по составу.
Характерной особенностью химического состава коры является высокое содержание экстрактивных веществ и наличие некоторых специфических компонентов, неудаляемых органическими растворителями. Массовая доля холоцеллюлозы в коре примерно в два раза меньше, чем в древесине, причем в лубе ее содержание выше, чем в корке. Целлюлоза в коре, как и в древесине, является основным полисахаридом, но в отличие от древесины она не является преобладающим компонентом коры. Массовая доля целлюлозы в коре 10-30 % [1].
Лигнин в тканях коры распределен менее равномерно, чем в древесине. Наружный слой коры более лигнифицирован, чем внутренний. Кора труднее делигнифицируется, чем древесина.
Бересту березы используют для получения дегтя. Пробковая ткань (дуб) служит для изготовления пробки. Из смоляных ходов коры пихты получают пихтовое масло. Из коры ели, дуба, ивы извлекают дубильные вещества. Кора может быть источником волокнистых полуфабрикатов для технических бумаг и картонов.
8. Физико-химические и физические свойства древесины
8.1. Влажность древесины и взаимодействие древесины с водой
Древесина обладает развитой капиллярной системой, и поэтому в древесине всегда содержится некоторое количество влаги.
Содержание воды характеризуется влажностью древесины. Различают два понятия: относительная влажность и абсолютная влажность.
Относительная влажность (влажность) – массовая доля воды, выраженная в процентах по отношению к массе влажной древесины.
Абсолютная влажность (влагосодержание) – массовая доля воды, выраженная в процентах по отношению к массе абсолютно сухой древесины.
Абсолютно сухой древесиной условно называют древесину, высушенную до постоянной массы при температуре 1050С (такая древесина содержит небольшое количество влаги).
Различают по влагосодержанию следующие виды древесины:
- свежесрубленная древесина с абсолютной влажностью 35-60 % (иногда до 100 %); зависит влагосодержание свежесрубленной древесины от времени рубки, породы древесины, условий произрастания;
- воздушно-сухая древесина – древесина, высушенная на воздухе до равновесного состояния ее влажности с относительной влажностью воздуха; абсолютная влажность такой древесины составляет 15-20 %;
- мокрая древесина – древесина после длительного нахождения в воде, с абсолютной влажностью выше 100 % (до 200 % и более);
- комнатно-сухая древесина – древесина, выдержанная в отапливаемом помещении, с абсолютной влажностью 8-12 %.
В процессе взаимодействия древесины с водой определяющее значение имеет гетерокапиллярная структура древесины. Большую роль при этом играют капилляры второго порядка (субмикроскопические капилляры). Они открываются при набухании древесины.
Древесина обладает способностью поглощать влагу из воздуха. Это ее свойство называется гигроскопичностью (влагопоглощением). При этом вода заполняет капилляры древесины второго порядка и адсорбируется поверхностями капилляров первого порядка; она называется гигроскопической или связанной влагой. При относительной влажности воздуха 100 % клеточной стенки полностью насыщаются влагой и достигают предела гигроскопичности. Этот предел называется точкой насыщения волокон. Равновесная абсолютная влажность в среднем составляет 25-30 %. Древесину, содержащую только гигроскопическую влагу, называют влажной древесиной. В таком волокне свободная влага еще отсутствует. При контакте с водой происходит дополнительное поглощение воды. Эта избыточная вода наполняет капилляры первого порядка (полости клеток) и рассматривается как свободная. Эта способность древесины впитывать воду называют водопоглощением, а такую древесину называют сырой древесиной.
Древесина и целлюлоза способны набухать в воде. Набухание увеличивает внутреннюю поверхность капилляров древесины и способствует дальнейшему поглощению воды. Максимум набухания достигается в точке насыщения волокна. Максимальная внутренняя поверхность набухшей древесины может достигать 200 м2/г. Обычно при этом линейное набухание не превышает 6-13 %. Затем поглощается свободная влага (до 200 % в зависимости от породы древесины и ее плотности).
Набухание сопровождается характерными для процесса набухания полимеров явлениями. При поглощении первых порций воды (до 6%) наблюдается контракция вследствие сжатия поглощенной воды примерно на 25 %. На этой стадии выделяется теплота набухания - теплота гидратации, зависящая от плотности упаковки компонентов древесины. Теплота гидратации древесины составляет 67-84 %. Влажность древесины и взаимодействие древесины и ее компонентов с водой имеет большое значение в технологии химической переработки древесины.
8.2. Физические и механические свойства древесины
Плотность древесины – величина, определяемая отношением массы древесины к ее объему и выражаемая в килограммах на кубический метр (кг/м3), в граммах на кубический сантиметр (г/см3). Она зависит от пористости и влажности древесины. Плотность древесины зависит также от породы и возраста древесины. Обычно плотность древесины ели составляет 0,37-0,58 г/см3, сосны 0,30-0,70 г/см3, березы 0,50-0,75 г/см3.
Для абсолютно-сухой древесины плотность и объем пор связаны следующей зависимостью:
r
С = 100 – (1 - ------),
d
где С – доля объема пор, % от объема древесины,
r - плотность древесины, г/см3,
d – истинная плотность вещества древесины, г/см3.
Истинная плотность вещества древесины приблизительно одинакова для разных пород (1,53-1,55 г/см3).
Теплопроводность древесины. Из-за пористого строения древесина плохой проводник тепла, что обусловлено присутствием в порах воздуха. Увеличение влажности повышает теплопроводность.
Удельная теплоемкость древесины практически зависит от плотности и древесной породы. Для абсолютно-сухой древесины при 00С ее значение 1,55 кДж/кг* 0С. С ростом температуры это значение увеличивается и при 100 0С оно больше почти на 25 %, чем при 00С.
Присутствие воды повышает теплоемкость древесины.
Абсолютно-сухая древесина имеет низкую электропроводность и соответственно высокое электрическое сопротивление (1014-1016 Ом*м).
Механические свойства отражают способность древесины сопротивляться действию усилий. К механическим свойствам относятся прочность и деформируемость. Они имеют большое значение при использовании древесины в качестве конструкционного материала.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК:
1. Азаров В.И., Буров А.В., Оболенская А.В. Химия древесины и синтетических полимеров: Учебник для вузов. СПб.: СПбЛТА, 1999. 628 с.
2. КулезневВ.Н., Шершнев В.А. Химия и физика полимеров. М.: Высшая школа, 1988. 312 с.
3. Никитин В.М., Оболенская А.В., Щеголев В.П. Химия древеесины и целлюлозы. М.: Лесная пром-сть, 1978. 368 с.
4. Оболенская А.В., Ельницкая З.П., Леонович А.А. Лабораторные работы по химиии древесины и целлюлозы. М.: экология, 1991. 320 с.
5. Боровиков А.М., Уголев Б.Н. Справочник по древесине. М.: Лесная пром-сть, 1989. 296 с.
6. Фенгел Д., Вегенер Г. древесина. Химия, ультраструктура, реакции: Пер.с англ. М.: Лесная пром-сть, 1988. 512 с.
7. Химия древесины/ Пер. с финского Р.В.Заводова под ред. М.А.Иванова. М.: Лесная пром-сть, 1982. 400 с.
8. Яковлев Г.П., Челомбитько В.А. Ботаника: Учеб. для фармац. Институтов и фармац. мед. вузов. М.: Высш.шк., 1990. 367 с.
Дата публикования: 2015-11-01; Прочитано: 1237 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
