 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Л1(17). ИСОН с высоким уровнем интеграции для высокоманевренных подвижных объектов
|
|
План
12.1 Окружающая среда
12.2 Условия среды
12.3 Организм и среда
12.4 Популяция и окружающая среда
12.5 Экосистемы
Каждый организм находится в многообразных взаимных связях с факторами окружающей среды, как абиотическими (геофизическими, геохимическими), так и биотическими (живыми организмами того же и других видов). Эти надорганизменные связи на различных уровнях интеграции изучает экология – наука о «хозяйстве» природы. Термин «экология» в 1866г предложил выдающийся немецкий биолог Э.Геккель. Изучаемые уровни – это взаимоотношения особей (аутэкология), популяций (популяционная экология или демэкология) и сообществ организмов (синэкология, системная экология) с их окружением.
12.1 Окружающая среда
К окружающей среде относятся те элементы среды, с которыми данный организм вступает в прямые или непрямые отношения. По характеру этих отношений окружающую среду можно разделить на разные «слои».
A. Воспринимаемая, информативная среда. Это специфичный для данного вида «фильтрат» окружающего мира. Человек не видит ультрафиолетовый свет, а пчела красный. Ультразвуковые локационные крики летучих мышей не воспринимаются человеком, а ночные бабочки прекрасно их слышат.
Б. Минимальная среда охватывает непосредственно необходимые для жизниэнергетические и материальные ресурсы окружающего мира (свет, пища, жизненное пространство и т.д.) независимо от того, все ли свойства этих ресурсов воспринимаются организмом. Например, существенно важное качество пищи – ее калорийность – не воспринимается органами чувств.
B. К физиологической среде относятся помимо указанных выше и другие, не необходимые для жизни, но все же действенныефакторы, как правило, в значительной мере определяющие оптимальную плотность популяций и ареалы: конкуренты, враги, паразиты, возбудители болезней, неблагоприятные крайние температуры, осмотическое давление и т.д.
Г. Экологическаясреда включает еще и те факторы, которые действуют непрямымобразом, влияя на минимальные и физиологические факторы. Например, для хищника выгодно, если хорошее пастбище повышает плотность популяции его жертв, и невыгодно, если популяция сокращается из-за эпизоотии или нехватки пищи.
В конечном счете, все организмы и факторы среды на Земле находятся в тесной или отдаленной связи между собой. Но так как земная поверхность дифференцирована, возникли более или менее разграниченные комплексы таких взаимоотношений. Определенные группы организмов так связаны потоками энергии и вещества, что образуют довольно стабильные во времени и пространстве надорганизменные образования – биомы: леса, степи, тундры и т.д. Для такого рода экосистем часто характерны определенные комбинации видов (сообщества организмов, биоценозы), а также определенные комплексы абиотических факторов среды.
Непосредственное окружение организма характеризуется с экологической точки зрения господствующими там абиотическими и биотическими факторами. Взятые в совокупности, они образуют местообитание организма, его «экологический адрес». Сравнение различных данных о местообитаниях какого-либо вида позволяет сделать выводы о факторах, относящихся к экологической среде данного вида и тем самым более или менее тесно привязывающих его к этим местообитаниям. Например, турбеллярия Planaria alpina встречается только в истоках горных ручьев; она приурочена к холодной, богатой кислородом воде. Для черного дятла необходимое условие – не леса с определенными видами деревьев, а наличие достаточно старых стволов, чтобы хватало мест для гнездования в дуплах. С другой стороны, многие виды приурочены к определенным местообитаниям. Например, сосны у нас произрастают на сухих, бедных питательными веществами почвах, так как в иных местах они не выдерживают конкуренции других деревьев, в свою очередь неспособных расти там, где могут расти сосны.
12.2 Условия среды
12.2.1 Общие геофизические условия в биосфере
К общим условиям существования живых организмов относятся наличие жидкой воды, ряда химических элементов (так называемых биогенных) и поступление лучистой энергии в диапазоне температур от –50 до +50 °С. Поэтому жизнь возможна только в тонком (и даже не везде непрерывном) слое между земной корой и атмосферой – в биосфере (рис 12.1).
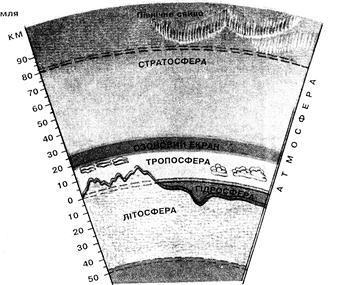
Рис. 12.1 Оболочки Земли
На высотах более 6000 м и в сухих областях длительное время способны выживать только покоящиеся стадии (например, споры); в темных глубинах вод и под землей могут жить лишь специализированные консументы и деструенты, используя биомассу, произведенную в освещенной зоне, или биомассу хемосинтезирующих бактерий. В океане освещенная зона редко заходит глубже 100 м, а в быстротекущих водах ограничена лишь несколькими сантиметрами. Хотя биомасса составляет по весу не больше 0,1% земной коры, в нее входят практически все элементы. Концентрация макроэлементов (С, Н, О, N, S, P, Ca, Mg, К, Na, Fe, CI) в биомассе превышает их концентрацию в земной коре, а микроэлементы (например, Al, Zn, Mn, Cu, Si, Br, I, As) обнаруживаются в биомассе обычно лишь в следовых количествах.
В энергетическом отношении жизнь в биосфере поддерживается постоянным притоком лучистой энергии от Солнца и использованием ее в процессах фотосинтеза. Этот процесс направлен против градиента энтропии, т.е. в живых организмах энтропия (в кажущемся противоречии со вторым законом термодинамики) уменьшается, но это происходит за счет ускоренного увеличения энтропии в окружающей среде.
Продуценты (растения, синезеленые водоросли, некоторые бактерии) осуществляют фотосинтез, при котором происходят эндергонические процессы образования молекул, богатых энергией (таких как углеводы или белки). Энергия света преобразуется при этом в химическую энергию синтезируемых молекул, а позже передается гетеротрофным консументам (прежде всего животным). В конце концов, не израсходованная на дыхание часть биомассы разлагается деструентами, также гетеротрофными (бактерии, грибы и др.). В итоге вся биомасса – иногда после длительной фоссилизации – при экзергоническом распаде высвобождает всю содержащуюся в ней энергию. Таким образом, экосистемы, хотя энергия в них на какое-то время задерживается, представляют собой энергетически открытые системы. Высвобождающаяся энергия безвозвратно теряется для системы (принцип энтропии!), а химические элементы могут использоваться снова в круговороте веществ.
Практически все вещества земной коры с разной скоростью и в разных количествах проходят через организмы. Существуют биогеохимические круговороты газового типа с очень протяженными в пространстве и очень подвижными резервуарами в атмосфере или океанах (циклы О2, СО2, Н2О, N2) (рис. 12.2, 12.3) и круговороты осадочного типа с менее протяженными резервуарами в земной коре (Fe, P, Са).
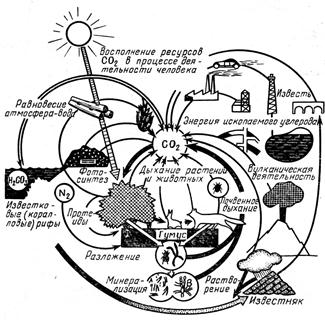
Рис. 12.2 Круговорот углерода в биосфере
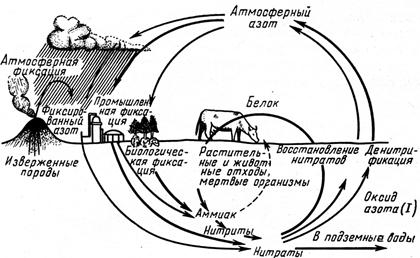
Рис. 12.3 Круговорот азота в биосфере
Помимо света, необходимого для фотосинтеза, решающие элементы климата, особенно для продуцентов, – это температура и влажность. Поток энергии, от которого зависят эти элементы, меняется от места к месту (зональные различия), на протяжении суток (суточные колебания) и на протяжении года (сезонные колебания). Эти явления связаны с вращением Земли и наклоном ее орбиты к плоскости вращения. На климатические циклы – суточные в тропиках и сезонные в умеренных поясах, влияет еще и расстояние до водных масс океанов (океанический или континентальный характер климата).
Метеорологи определяют местный климат по измерениям, проводимым в метеобудках на высоте 2 м над поверхностью земли. Но в различных биотопах, прежде всего вблизи почвы, условия могут быть иными, особенно в отношении баланса излучения.
Из достигающего земной атмосферы солнечного излучения, в основном коротковолнового (8,12 Дж·см-2∙мин-1 – это, так называемая, солнечная постоянная), днем в атмосфере 42% отражается (33% – отражение от облаков, 9% – диффузное отражение) и 15% поглощается (рис. 12.4). Только остальные 43% (так называемая суммарная радиация) достигают почвы (27% прямо и 16% в виде рассеянного света неба даже при сплошной облачности). Только часть этой энергии проникает в глубь почвы или воды путем теплопроводности. Остальная энергия расходуется на конвекцию воздуха и парообразование, отражается (коротковолновая часть спектра) или переизлучается. Часть излученной энергии возвращается на почву в результате рассеяния атмосферой (водяным паром, СО2, озоном) в виде длинноволнового противоизлучения(тепла). Размеры потерь энергии в значительной степени зависят от свойств почвы или воды (от плотности, влажности и цвета почвы, от наличия снега, от высоты и плотности растительного покрова и т.п.).
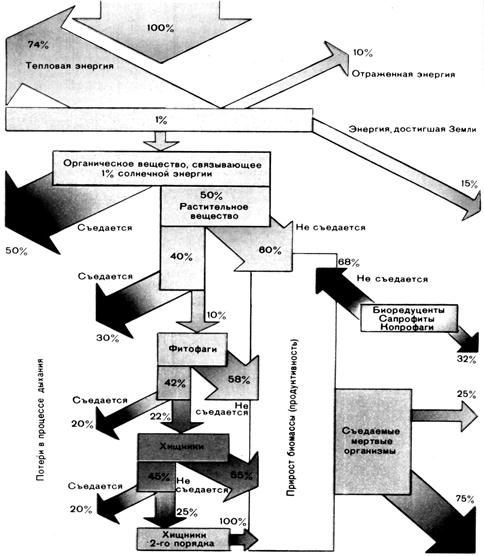
Рис. 12.4 Поток энергии через лиственный лес
На экоклимат сильно влияет растительность. При плотном растительном покрове (поля, луга, леса) интенсивность лучевого потока на его верхней границе такая же, как на голой почве, а у основания – часто более чем в 10 раз меньше. Поэтому наземная растительность густого букового леса ассимилирует в основном до появления листвы на деревьях. Поскольку древесный полог тормозит также излучение и испарение из глубины растительного покрова, там складывается ровный микроклимат с малыми амплитудами колебания температуры, повышенной влажностью воздуха и ослабленным его движением (рис. 12.5).

| 
| 
| 
| 
|
| Растительность | Температура | Свет | Скорость ветра | Относительная влажность воздуха |
Рис. 12.5. Микроклимат в плотной растительности (сильно схематизировано). (По данным Geiger)
Другие различия в экоклимате определяются топографиейместности. Южный склон при сильных различиях инсоляции и излучения в общем теплее, суше и имеет более длинный световой день, чем северный склон. Наклон местности облегчает стекание холодного воздуха и влияет на инсоляцию. Так возникают экоклиматические островки, где могут жить организмы, уже неспособные существовать в окружающей местности. Например, в Средней Европе на северных склонах сохраняются северные или альпийские виды, а на южных склонах – виды, свойственные южной и юго-восточной Европе.
12.2.2 Особенности субстрата
Так как процессы обмена веществ могут идти только в водных растворах, организмы вначале смогли заселить одну лишь гидросферу. Позже они вышли и на сушу в тех местах, где есть достаточно воды, а в свободном воздушном пространстве организмы могут находиться лишь временно (при добывании пищи, расселении и т.п.).
В более или менее крупных водоемах есть два местообитания: свободная водная масса (пелагиаль) и донная зона (бенталь). И та и другая подразделяются на нижний, слабо освещенный этаж, где нет фотосинтеза, и верхний, пронизываемый светом. Обитатели пелагиали предотвращают свое погружение на глубину либо активным плаванием, либо пассивно – с помощью приспособлений для парения в воде. Сильные пловцы (нектон) передвигаются собственными усилиями, а остальные – планктон – переносятся течениями. Многие из прикрепленных или ползающих обитателей бентали часть жизни проводят как планктонные организмы, по крайней мере, на личиночных стадиях.
Для доставки кислорода в глубинные слои, а питательных веществ – в верхние решающее значение имеет перемешивание воды. Самые рыбные районы океана находятся там, где морские течения, выносят в пелагиаль из глубинных осадочных резервуаров достаточные количества неорганических питательных веществ (особенно соединений N и Р).
Для наземных экосистем решающее значение имеет доступность воды. Пресная вода составляет всего лишь около 3% общего запаса воды на Земле (1300 млн. км3). Примерно 3/4 имеющейся пресной воды лежит в виде льда, а в атмосфере циркулирует только 0,35%. При этом 78% испарившейся воды циркулирует над морем, 14% над сушей и только 8% между морем и сушей. Большая часть воды атмосферных осадков сразу задерживается в растительности (например, в травостое клевера 40%, во мхе и подстилке 18%), так что в почву попадает только 1/3 –1/4 выпавших осадков. Эвапорация (испарение со свободной поверхности) и транспирация (испарение растительными тканями) возвращают воду в атмосферу, причем площади, занятые растительностью, особенно леса, отдают больше, чем голые участки. Вода осадков, не стекшая по поверхности в какой-либо водоем, просачивается в виде гравитационной воды в пустоты почвы, где она в большей или меньшей степени удерживается в капиллярах, как пленочная вода (водяная пленка вокруг частиц почвы) или силами когезии как капельножидкая вода (почвенная влага), а частично может проникать в грунтовые воды. Кроме грунтовых вод, до которых добираются лишь некоторые корни, в распоряжении растений находится только капиллярная вода, висящая в капиллярах и временно наполняющая пустоты.
Почва – самый наружный, разрыхленный физическим и химическим выветриванием слой земной коры – является одним из самых важных субстратов жизни. Множество организмов, в основном мелких (так называемый эдафон), обитает в почве более или менее постоянно (рис. 12.6).
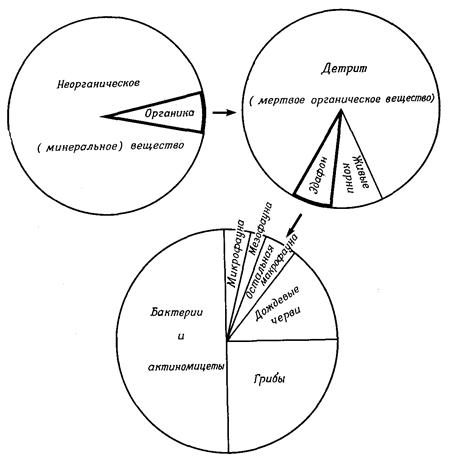
Рис. 12.6. Общий состав верхнего слоя почвы и его эдафона (по весу) в смешанном лесу. (По Dunger, с изменениями)
Эти организмы вносят решающий вклад в образование почвы, механически разрыхляя и перемешивая ее (дождевые черви), способствуя растворению определенных веществ (с освобождением СО2), создавая запасы отмершей биомассы и продуктов ее разложения и реминерализации, в том числе гуминовых веществ, питательного гумуса и не поддающегося дальнейшему разложению стойкого гумуса - резервуаров неорганических питательных веществ для высших растений. Поэтому состав и структура почв – это один из важнейших факторов, определяющих само существование как низших, так и высших растений (а также распространение и образование сообществ).
12.3 Организм и среда
Каждый организм обладает способностью реагировать в соответствии со своей генетической конституцией на окружающую среду, использовать ее факторы для своего существования и развития или, по крайней мере, переносить их воздействие. Эта экологическая потенция определяется наследственной нормой реакции по отношению к каждому фактору среды. Норма реакции у каждого вида характеризуется определенным положением и диапазоном (широтой) на шкале интенсивности данного фактора – экологической валентности. Экологическая потенция охватывает диапазон от нижнего пессимального предела (минимума) до верхнего пессимального предела (максимума) с оптимумом где-то посередине (рис. 12.7). В пессимумах способность реагировать снижается до нуля, за ними наступает абсолютное торможение процесса (реакции), часто даже необратимое повреждение организма. Широта экологической потенции (от минимума до максимума) у эврипотентныхвидов велика, у стенопотентныхмала. Смотря по тому, где лежит оптимум для стенопотентного вида — в нижней, средней или верхней части всего диапазона, — различают олиго-, мезо- и полипотентные виды. Положение иширота нормы реакции зависят от возраста, пола и фазы развития и различны для разных процессов (активности, метаболизма, развития и т.д.). Границы экологической потенции могут (обычно лишь ненамного) сдвигаться в результате модификации, т.е. индивидуального приспособления (привыкания) или мутации, т.е. генетического приспособления
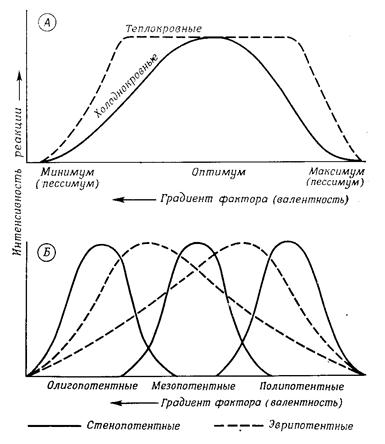
Рис. 12.7. Экологическая потенция в сопоставлении с областью валентности фактора среды А. Общая схема для пойкилотермных и гомойотермных животных. Б. Различные положения и ширина области потенции (норма реакции)
12.3.1 Фактор температуры
Температура влияет на энергетику всех жизненных процессов. Так как в основе всех реакций живого организма, зависящих от температуры, в конечном счете лежат биохимические процессы, для них в основном верно правило зависимости скорости реакции от температуры – закон Вант-Гоффа, согласно которому при повышении температуры на 10°С реакция ускоряется в 2-3 раза. Однако под влиянием эндогенной нормы реакции и лимитирующих внешних факторов соответствующая экспоненциальная кривая рано или поздно переходит в кривую с оптимумом.
Активность животных ограничивается пессимумами, при которых наступает обратимое тепловое или холодовое окоченение. У насекомых повышение температуры вызывает вначале медленные, некоординированные движения, в физиологической области (оптимум) приводит к полностью управляемой активности, а при дальнейшем повышении – к чрезмерно быстрым, некоординированным, суматошным движениям.
Главная точка приложения для воздействия температуры – обмен веществ. Для ассимиляции, как и для диссимиляции, существуют специфические температурные пределы и специфический оптимум.
Эвритермныеорганизмы (с широкой экологической потенцией) – это, например, актиния Actinia equina, которая может жить как в тропических, так и в арктических морях; карпы, способные жить и в холодных, и в теплых пресных водоемах. Стенотермныевиды приспособлены к узкому диапазону температур. Среди них политермны (т.е. приспособлены к теплу) почти все рифообразующие кораллы, термиты, тараканы идругие животные; олиготермны(приспособлены к холоду) гренландский кит, самцы зимней пяденицы, ручьевая форель, сардиновые рыбы.
Подвижные организмы могут использовать внешние источники тепла или избегать их, изменяя позупо отношению к источнику теплового излучения (гелиорегуляция) или отыскивая места, более подходящие в отношении тепла (термотаксис). Холодным утром кузнечики подставляют бока солнечному свету, а дневные бабочки расправляют крылья; в полуденную жару они, сложив крылья, располагаются параллельно лучам. Высокие компасные термитники, так же как листья компасных растений, ориентированы широкими сторонами на восток и запад (что дает выигрыш в инсоляции утром и вечером), а узкими – на север и юг (так что нет перегрева в середине дня).
Термопреферендум(предпочтительная температура) специфичен для каждого вида; это та температура, при которой не расходуется энергия ни для производства тепла, ни для его отдачи. Организм обычно находит такие условия путем фоботаксиса, в результате беспорядочных передвижений (методом проб и ошибок).
Предпочитаемая температура (ПТ), в общем, соответствует экоклимату нормального местообитания данного вида. У олиготермной форели (из верховий текучих вод) она составляет 10,5°С, у эвритермных карпов (стоячие воды) – 21,3. Она может быть различной на разных стадиях развития. Строительство гнездслужит у наземных беспозвоночных, птиц и млекопитающих прежде всего для сохранения тепла, выработанного в организме или полученного от внешнего источника. Общественные пчелы летом поддерживают в улье удивительно постоянную температуру – от 34,5 до 35,5°С, согревая его движением крыльев или мышц либо охлаждая вентиляцией и испарением принесенной воды.
Только гомойотермные (теплокровные) животные-птицы и млекопитающие – сравнительно независимы от окружающей температуры и создают свой собственный «внутренний климат», поддерживая постоянную температуру тела (37-39°С у млекопитающих, 41-42°С у птиц). Поэтому кривые их реакции на температуру образуют почти ровное плато между пессимумами (рис. 12.7). И все же у этих животных тоже есть предпочитаемые температуры (такие, при которых им приходится тратить минимум энергии на терморегуляцию); они обладают такими приспособлениями, как сезонные изменения густоты и длины шерсти, а для выращивания молоди используют теплоизолирующие материалы. Поэтому многие из них могут жить почти во всех климатических зонах (например, пума и тигр – от субарктики до тропиков).
При сравнении родственных видов из теплых и холодных областей часто можно отметить увеличение размеров тела (правило Бергмана) или уменьшение придатков (конечностей, ушей, хвоста; правило Аллена) от теплых районов к холодным.
12.3.2 Водный режим
Обменные процессы - биохимические реакции, которые протекают в водных растворах. Поэтому для активной жизни необходимо достаточное содержание воды в организме (у наземных животных 45-95% веса тела). Так как давление водяного пара в воздухе, как правило, сравнительно невелико, неизбежны потери воды в результате транспирации, а покрыть все тело водонепроницаемой изоляцией (кутикулой, слоем воска, роговым панцирем) невозможно из-за необходимости газообмена при дыхании и фотосинтезе. Поэтому запас воды приходится часто пополнять путем питья (позвоночные, пауки, брюхоногие моллюски) или потребления влажной пищи (виды, питающиеся листьями и плодами). Обитатели аридных районов используют метаболическую воду, образующуюся при окислении жиров и других веществ; жир запасается, например, в горбах верблюдов. При высокой влажности воздуха возможно поглощение влаги через покровы тела (амфибии, брюхоногие моллюски, тропические растения-эпифиты), иногда за счет гигроскопичности каких-либо структур – против градиента влажности (мокрицы, клещи, мучной хрущак). Растения аридных местообитаний (пустынь) обладают очень широко расходящимися или очень глубоко проникающими в землю корнями с низким (сильно отрицательным) осмотическим потенциалом.
Потери на транспирацию уменьшаются благодаря защитным покровам и подушкам неподвижного воздуха (удерживаемого шерстью и перьями у животных, волосками у растений), обратному всасыванию воды при выделении мочи и дефекации, поискам влажных мест. Муравьи, термиты, комары активно разыскивают подходящие слои воздуха (гигротаксис). Большинству видов свойственно фоботаксическое (гигрокинетическое) поведение: их ненаправленная активность успокаивается только в месте с подходящей влажностью воздуха. Гигрофильныевиды почти не имеют защиты от высыхания и могут существовать только в очень влажном воздухе (амфибии, безраковинные брюхоногие моллюски, большинство обитателей почвы, травы тропического дождевого леса). Ксерофильныевиды обладают либо морфологическими средствами защиты от высыхания, либо могут накапливать воду (пустынные суккуленты, например кактусы), всасывать ее обратно, в сухое время суток разыскивать укрытия или восполнять потери воды большим количеством влажной пищи (рис. 12.8).

Рис. 12.8. Растения, принадлежащие к различным экологическим группам
12.3.3 Фактор света
Являясь источником энергии для фотосинтеза, свет имеет фундаментальное экологическое значение. Даже более или менее лишенные света экосистемы глубинных морских вод, почв и пещер в конечном счете зависят от первичной продукции биомассы зеленых растений на свету.
L – освещенность в данном местообитании по сравнению с полной освещенностью непокрытой почвы. Светолюбивые растения пустынь, степей, тундр и высокогорья могут ассимилировать только при L = 100%. У теневыносливых растений точка компенсации (при которой ассимиляция СО2 равна высвобождению СО2) достигается при гораздо меньшем световом довольствии, а неослабленный свет им вреден (рис. 12.9). Луговые растения занимают среднее положение. На состав растительного покрова сильно влияют и видоспецифические требования растений к освещенности, и местные условия освещения. Лесной бук, теневыносливый и в молодом возрасте, вытесняет светолюбивые дубы и сосны; под пологом его крон могут существовать только теневые растения.
Свет вызывает в органах растений (стеблях и их разветвлениях, черешках листьев и т.д.) движения типа искривлений, направленные на оптимальное использование света листьями, т.е. наивысшую активность фотосинтеза (фототропизм).
Так называемым растениям длинного дня (в основном это растения средних широт или происходящие оттуда) для начала цветения требуется, в общем, более длительный светлый период суток (>12-14 ч в сутки), чем растениям короткого дня, происходящим из экваториальных областей (<12 ч в сутки), поэтому последние могут у нас цвести только осенью (или весной).
12.3.4 Экологическая потенция
По экологической потенции в отношении температуры, света, влажности воздуха, содержания солей и кислорода в воде и т.д. различают виды с широкой и узкой нормой реакции: эвритермные и стенотермные, эвригидрические и стеногидрические эвригалинные и стеногалинные, эвриоксибионтные и стено-оксибионтные. Лишь у немногих видов требования ко всем илибольшинству факторов среды так малы, а широта реакции соответственно так велика, что они могут существовать практически во всех местообитаниях данной области, то есть быть эвриойкными.Полностью эвриойкные виды и настоящие космополиты редки (человек, крыса). Стеноойкныевиды специализированы для жизни в определенных местообитаниях; но эврипотентные виды тоже могут оказаться стеноойкными в тех областях, где их требования, например к температуре, удовлетворяются только в отдельных специфических местообитаниях.
В центре ареала данного вида диапазон его потенции часто в значительной мере совпадает с широтой валентности большинства факторов среды, а на окраинах ареала, напротив, тот или иной фактор часто приходится на область пессимума.
Редко бывает так, чтобы в одном месте области валентности всехжизненно важных факторов совпадали с диапазонами потенций вида. Чаще всего хотя бы один фактор лежит вне оптимума. Тогда от этого фактора зависит возможность существования вида в данном месте. Экологически решающим всегда оказывается фактор, оказавшийся пессимальным для той стадии, которая обладает наименьшей широтой реакции.
Выход за пределы пессимума не всегда опасен для индивидуума. Возможны два способа «эмансипации» от экологической валентности: сдвиг диапазона потенции путем акклиматизации(например, спячка) и уклонение от неблагоприятного фактора среды посредством миграции.
Успешность акклиматизации к низким температурам (резистентность к холоду) у пойкилотермных (холоднокровных) животных зависит от предшествующего опыта. У гомойотермных животных снижение температуры действует не только закаливающим образом, но и способствует образованию подкожных жировых отложений и более густого меха.
У холоднокровных и примитивных теплокровных животных пессимальные условия среды вызывают спячку, т.е. прекращение активности и замедление процессов метаболизма и развития, немедленно и неизбежно. Гораздо чаще спячка наступает лишь после более длительного воздействия неблагоприятных условий – олигопаузы, часто на вполне определенной стадии (на стадии яйца, личинки, куколки, у взрослого животного) (рис. 12.9). При этом начало и окончание паузы часто определяются уже не одним только пессимальным фактором, а в основном длиной светового дня (фотопериодом). Длина дня, регулярно изменяющаяся на протяжении года, предвещает приближение благоприятных и неблагоприятных сезонов точнее, чем все другие, менее регулярные колебания климатических факторов.
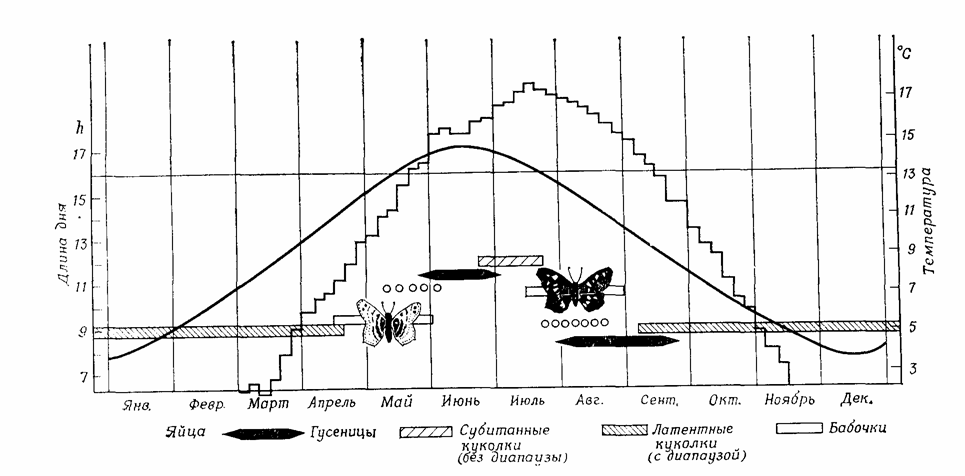

Рис. 12.9. Фотопериодическая индукция диапаузы:возникновение сезонных форм Araschnia levana в результате развития куколок с диапаузой или без нее. Вариант развития определяется длиной светового дня на стадии гусеницы. Ступенчатая линия – средние температуры за пятидневку. Плавная кривая – длина дня (включая «гражданские сумерки»). Горизонтальная линия – критический фотопериод. (По Miiller, с изменениями.)
Ухудшение условий среды может также приводить к миграциям, причем последние иногда совершаются заблаговременно. Недостаток пищи или ухудшение погоды побуждает некоторых летающих насекомых (саранчу), птиц (клеста, кедровку, свиристель), а также млекопитающих (леммингов) к первоначально ненаправленным откочевкам. Регулярные миграциимногих перелетных птиц определяются ежегодно изменениями погоды (так называемые погодные птицы – грачи, лысухи, зяблики, дрозды и др.). Другие («инстинктивные») птицы отлетают в более или менее фиксированные сроки, руководствуясь, по-видимому, фотопериодом, задолго до ухудшения погоды и сокращения пищевых ресурсов (таковы, например, кукушка, черные стрижи, аисты, иволга).
12.4 Популяция и окружающая среда
Решающие биотические элементы естественных экосистем – это не особи, а популяции. На их структуру и динамику влияют не только абиотические, но, прежде всего, биотические факторы. Плотность популяции (обилие), т.е. число (или биомасса) особей на единицу площади или объема, возрастной состав и генофонд – основные параметры экологии популяций.
Плотность популяции максимальна, когда она такова, что большая плотность уже не могла бы поддерживаться данной экосистемой, и минимальна, когда при меньшей плотности члены популяции уже не имели бы нормальных шансов на размножение (на встречу партнера).
12.4.1. Изменения плотности популяции
Если не учитывать иммиграцию и эмиграцию, плотность популяции определяется внутренними (конструктивными) факторами – рождаемостью и смертностью, а на них в свою очередь влияют внешние (регуляторные) абиотические и биотические факторы.
Общая рождаемость– это число новых особей, (D Nn), добавляющихся в единицу времени (D t). Так как оно зависит от числа уже имеющихся особей, лучше использовать удельную рождаемость:
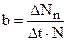 (12.1)
(12.1)
Идеальная(максимально возможная) рождаемость реализуется только в оптимальных условиях. Сопротивление среды (нехватка пищи, партнеров, мест для размножения, неблагоприятная погода и т.п.) сводит идеальную рождаемость к реальной(экологической).
То же, но с обратным знаком верно и для смертности. Это число особей, погибающих в единицу времени (D Nm). Удельная смертность (d) получится, если отнести эту величину к численности популяции:
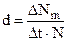 (12.2)
(12.2)
Идеальная(минимальная) смертность проявляется как результат физиологической смертности от старости только при оптимальных условиях жизни. Сопротивление среды (погода, конкуренция, враги) повышает ее до реальной(экологической) смертности.
От баланса между рождаемостью и смертностью (b > < d) зависит, в какой мере изменяется плотность популяции. В идеальных условиях (при максимальной рождаемости, минимальной смертности и стабильном возрастном составе) скорость роста популяции r1=b-d всегда принимает положительные значения (b>d), так как организмы всегда дают больше способного к размножению потомства, чем было бы необходимо, чтобы заменить родителей. Поэтому популяция непрерывно растет. Специфичный для каждого вида репродуктивный потенциал(скорость размножения, биологический потенциал, или потенциальная сила размножения)
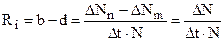 (12.3)
(12.3)
при полном отсутствии сопротивления среды дает экспоненциальный рост популяции.
В природных условиях рост популяции рано или поздно прекращается из-за сопротивления среды,которое увеличивается уже из-за того, что возрастает плотность популяции. Поэтому реальная кривая ростаобычно принимает сигмовидную (логистическую) форму.
После начальной логарифмической фазы она асимптотически приближается к уровню максимальной плотности популяции, т.е. к плотности насыщения (емкости среды - К), причем b становится равным d (рис. 12.10).
Размер популяции поддерживается на уровне К разными способами. У видов, живущих в эфемерных местообитаниях с высоким сопротивлением среды (большие потери от врагов), или у паразитов (малые шансы найти хозяина) репродуктивный потенциал п должен быть очень большим (r1 -стратеги), чтобы они могли быстро использовать редкие или небольшие шансы. Напротив, виды, живущие в долговременных, стабильных местообитаниях, с небольшим количеством врагов или совсем не имеющие их, виды с развитой заботой о потомстве, образующие семьи или
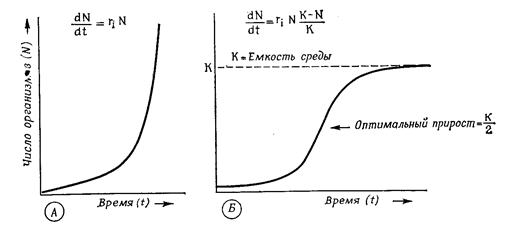
Рис. 12.10. Рост популяции. А. Экспоненциальная кривая роста при идеальных условиях среды. Б. Логистическая кривая роста в естественных условиях при емкости среды, равной К. (Wilson, Bossert)
стада, обходятся небольшим репродуктивным потенциалом (К -стратеги), так как у них значительная часть потомков достигает репродуктивного возраста. Например, орлы, дельфины и крупные копытные дают в год лишь одного потомка.
12.4.2 Влияние биологических факторов
В регулировании плотности популяций играют значительную роль внутривидовые и межвидовые взаимодействия. Особенно велика роль конкуренции за ресурсы.
Внутривидовая конкуренция (за пищу, полового партнера, жизненное пространство, место для размножения) увеличивается с ростом плотности популяции и степени специализации вида. Чаще всего начинается конкуренция за пищу, когда в результате размножения при еще достаточном запасе пищи плотность популяции повышается. Недостаточное питание может тогда приводить, например, к снижению плодовитости, пока уменьшение популяции не позволит виду снова размножаться.
Даже при достаточном количестве пищи высокая плотность – фактор скученности – может снижать плодовитость. У дрозофилы, например, это происходит в результате нарушения брачного поведения, взаимных помех между яйцекладущими самками, питающимися личинками и покоящимися куколками. Высокая плотность иногда приводит к каннибализму даже у видов в норме чисто растительноядных.
Организмы противодействуют слишком высокой плотности популяции по так называемому принципу конкурентного исключения. Активное противодействие принимает форму территориального поведения, пассивное состоит в дисперсии (рассеянии), т.е. разрежении популяции путем более равномерного распределения в пространстве.
Территориальность – явление, основанное на врожденном стремлении особи к свободе передвижения на некоторой минимальной площади. Первая ступень развития территориальности – индивидуальное пространство, окружающее каждую особь (оно хорошо заметно у ласточек, усевшихся на телефонный провод, или у скворцов в летящей стае). Особь защищает его от вторжения и открывает для другой особи только после церемоний ухаживания перед спариванием. Вторая ступень – обороняемое место для жизни, отдыха или сна в середине необороняемой зоны активности (у многих хищников – охотничьего участка). Самое рациональное использование пространства достигается на третьей ступени территориальности, когда образуются настоящие территории – участки, из которых другие особи изгоняются.
Межвидовые взаимоотношения могут быть безразличными, вредными или полезными для партнеров. При нейтрализмеоба вида живут в одной экосистеме, не вступая в отношения друг с другом (например, гидроидные полипы на раковине моллюска, дятлы неподалеку от дроздов в буковом лесу). Может существовать конкуренцияза одинаковую пищу или жизненное пространство (например, между корнями деревьев в саванне, между полевыми воробьями и синицами за места гнездования). Мутуализмприносит выгоду обоим партнерам – при симбиозежизненно важную, при протокооперации– не очень значительную (рис. 12.11). Жвачные животные и микроорганизмы их рубца не могут существовать друг без друга; напротив, гидра может жить без водоросли хлореллы, как и та без нее.

| 
| |
| Актиния на панцире рака отшельника | Лишайник | |

| 
| |
| Симбиоз орхидеи и гриба. А- цветки северной орхидеи-ятрышника. Б – гифы гриба в клетках ятрышника, В – проросток орхидеи с участком, занятым мицелием | Схема связи мицелия гриба с корнями дерева – микоризы | |
Рис. 12.10 Симбиоз
Чаще польза и вред бывают односторонними. Для льва безразлично, поедают ли грифы и шакалы остатки его трапезы (комменсализм) (рис. 12.11);для жуков-навозников несущественно, что в полете они переносят нематод-копрофагов к новым навозным кучам — их субстрату (форезия).При паразитизмеи хищничествеодин из партнеров извлекает для себя пользу во вред другому. Эти два типа взаимоотношений различаются тем, что в первом случае нападающий организм меньше своей жертвы, аво втором — крупнее. Кроме того, репродуктивный потенциал у паразита больше, чем у хозяина, а у хищника — меньше, чем у жертвы.

| 
|
| Орхидея, которая поселилась на стволе дерева | Средиземноморский краб, который носит в клешнях актинии |
Рис. 12.11. Примеры комменсализма
В гомогенной среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам. В естественных условиях возникает следующая временная и причинно - следственная цепь: размножение жертвы → размножение хищника → резкое сокращение численности жертвы → падение численности хищника → размножение жертвы и т.д. (рис. 12.12).Эта кибернетическая система с отрицательной обратной связьюприводит к устойчивому равновесию; волны флуктуации хищника и жертвы следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника, так и жертвы остается постоянной.
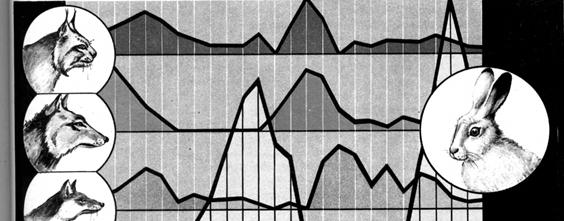
Рис. 12.12. Связь численности хищников (рысь, волк, лисица) и жертвы (зайца) в центральных районах европейской части России
Для популяции жертвы влияние хищника положительно, так как чрезмерное размножение жертвы привело бы снова к резкому падению ее численности. К тому же хищник в основном истребляет больных и слабых особей, которые действуют как фертильные внутривидовые конкуренты. В свою очередь все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.
Защита от врагов может быть активной – например, укусы, уколы, удары (в том числе электрические – у электрических скатов, сомов и других рыб), выбрызгивание секретов и т.п., использование укрытий – или, гораздо чаще, пассивной, к которой относятся маскирующая (мимикрическая) внешность, предостерегающая внешность (мимикрия в широком смысле слова), маскирующее или предостерегающее поведение. У растений развиваются колючки, шипы, жгучие волоски, яды, горькие вещества.
Маскирующая внешностьсостоит в подражании несъедобным предметам (например, палочники и гусеницы пядениц имитируют сучки, палочник «бродячий лист» и отдыхающие бабочки – листья, долгоносик Llthinus – лишайники) или зрительном слиянии с окружающим фоном: зеленая окраска обитателей листвы (квакши, кузнечики, клопы, гусеницы), коричневая – у наземных обитателей (жаворонки, песочники, самки уток). Иногда приспособление к цвету и узору субстрата может осуществляться путем физиологического изменения окраски тела (каракатицы, скаты, камбалы, квакши) или переменой окраски при очередной линьке (кузнечики).
Предостерегающая внешность может использоваться с двумя целями. Одна цель – отпугивание агрессора необычным рисунком, глазчатыми пятнами, появляющимися у многих бабочек, когда они раскрывают крылья, имитацией змеиной головы (у многих гусениц) или имитацией животных, опасных для нападающего (отпугивающая внешность). Другая возможность – предупреждение яркими сигнальными цветами и бросающимся в глаза рисунком о реальных отрицательных для нападающего свойствах жертвы: горьком вкусе, несъедобности, ядовитости, умении кусать или жалить. Примеры – божьи коровки, клопы-арлекины, пестрые гусеницы, осы. Но при этом приносится в жертву какая-то часть популяции, на которой агрессор усваивает горький опыт. Некоторые безвредные организмы имитируют предупреждающую окраску опасных видов (например, мухи-журчалки, многие жуки-усачи и бабочки-бражники подражают внешности ос). Они, можно сказать, пользуются защитой своих образцов, копируя их: эта мимикрия в узком смысле слова – обман.
12.4.3 Регулирование плотности популяции
Рост популяции регулируется факторами, зависящими и не зависящими от ее плотности.
Не зависящие от плотности факторы, такие как большинство климатических факторов (температура, влажность воздуха, количество осадков и др.), ускоряют или тормозят рост популяции всегда в степени, пропорциональной исходной плотности. Так, например, наступление холодов снизило количество рыбы в трех прудах одинаково на 6-8%, хотя плотность популяций была очень разной (от 55 200 до 2000 рыб на один день ловли) и осталась разной. То же можно сказать и о благоприятных погодных условиях: они могут вызывать массовое размножение независимо от того, высокой или низкой была исходная плотность.
В отличие от этого факторы, зависящие от плотности, как, например, большинство биотических факторов (конкуренция, враги, паразиты, патогенны), влияют на рост популяции только лимитирующим образом, и их действие никогда не бывает одинаковым при разной плотности популяции. Они действуют как регуляторы в узком смысле слова, «подгоняя» популяцию к существующей в данный момент емкости среды. В плотной популяции выше вероятность контактов и распространения инфекций. Отдельный хищник сокращает плотную популяцию жертв меньше, чем редкую, но только в том случае, если он не размножается. Таким образом, действие факторов, зависящих от плотности, сильно зависит также от общей ситуации в экосистеме.
12.5 Экосистемы
Экосистемы – это надорганизменные объединения с определенным видовым составом. В экосистемах суши (лесах, степях, лугах, болотах и т.д.), прежде всего, бросаются в глаза неподвижные, обычно хорошо заметные и обладающие большой биомассой продуценты. Состав растительных сообществ в экосистемах характеризуется, однако, не систематическими категориями, а жизненными формами. Переходы между экосистемами часто постепенны (например, между лесом и степью), границы их нечетки (например, между топяным болотом, тростниковым болотом и озером). Будучи энергетически и структурно открытыми системами,они обязаны своим единством взаимодействию своих компонентов, которые поддерживают в отношении друг к другу и кнеживой среде статистическое (подвижное) равновесие. Поэтому экосистемы характеризуются не столько определенными числами и неподвижными границами, сколько средними значениями и границами разброса.
12.5.1 Структура экосистем
Из многих сходных по своей биологии видов одной экосистемы (например, насекомых, сосущих соки луговых растений, – тлей, цикад, клопов) обычно лишь немногие (5-10%) составляют основную часть (около 4/5 ) биомассы данной группы. Высокая плотность популяций этих доминирующих видов говорит об их оптимальной приспособленности к данной экосистеме и большом для нее значении. Часто они встречаются и в других экосистемах, но образуют там популяции меньшей (субдоминанты) или совершенно незначительной (рецеденты) плотности. В одинаковых экосистемах удаленных друг от друга областей один вид может быть заменен другим: например, в американских тропических лесах среди птиц опылителями цветков служат колибри, ав африканских – медососы. Поэтому экосистему редко можно охарактеризовать перечнем отдельных видов или, в лучшем случае, комбинацией видов. Для этого больше всего пригодны стеноойкные виды, приуроченные к особым условиям данной экосистемы; изредка это бывают доминантные виды (средний дятел в лиственном лесу, клюква на торфяном болоте).
Для вида как структурного элемента экосистемы решающее значение имеет не его систематическое положение (отражающее гомологии в его плане строения), а функциональная конституция и экологическая потенция. Поэтому вид представляет собой комплекс жизненных форм, которые позволяют ему использовать определенные свойства (экологические лицензии) среды (субстраты, источники пищи, диапазоны температур и влажности и т.д.) и играть таким образом определенную роль в экосистеме.
Как один вид может образовывать целый спектр жизненных форм, так каждое местообитание предоставляет организму целый спектр факторов в виде экологических лицензийдля использования. Из того и другого – из сочетания области потенции определенной жизненной формы и области валентности необходимых для ее реализации факторов среды – при достаточном совпадении складывается экологическая ниша.Жизненную форму можно сравнить с профессией, имеющуюся валентность – с рабочим местом, а экологическую нишу – с реализованными трудовыми отношениями. Говоря об экологической нише вида, имеют в виду всю многомерную мозаику ниш. И наоборот, каждое местообитание постоянно предоставляет лицензии на ниши для множества организмов. Здесь господствует принцип конкурентного исключения, допускающий в каждую нишу (в узкомсмысле слова) только один вид.
Возможность внедрения случайно завезенных (колорадский жук) или самопроизвольно распространившихся (кольчатая горлица) видов показывает, что в экосистеме – по историческим и экофаунистическим или экофлористическим причинам – еще могут быть свободные, долго не используемые лицензии. Это может быть связано, например, с тем, что или в область вторжения нового вида еще не проникли противодействующие ему виды, или они были, но исчезли.
Совокупность всех видов, которые встречаются совместно, называют сообществом организмов, или биоценозом, а занятое ими местообитание, т.е. совокупность всех экологических факторов их местонахождения, – биотопом(«место жизни»). Но поскольку биоценоз трудно четко отделить от биотопа, лучше объединять их под именем биогеоценоза (рис. 12.13).Биогеоценозу свойственно общее энергетическое и материальное «хозяйство», более или менее отграниченное от других аналогичных комплексов, а также способность к самоподдержанию (путем регуляции).

Рис. 12.13. Биогеоценоз леса (показаны энергетический вход и выход)
Большинство биогеоценозов подразделяется на субъединицы (ярусы, биохоры, меротопы), которые увеличивают возможное число ниш и видов, но в то же время экологически взаимозависимы.
Наиболее стабильны и замкнуты ярусы («этажи»), например, в лесу –древесный ярус, кустарниковый, травянистый и напочвенный. Хотя каждый из них обладает собственным микроклиматом, а часто и собственной фауной (тли, короеды), они связаны между собой климатически и функционально (например, пищевыми цепями с участием листового опада). Многие птицы, гнездящиеся на земле (пеночки и др.), разыскивают себе пищу в кустарниковом и древесном ярусах, а гнездящиеся в кроне (голуби) – на почве.
В менее самостоятельных биохорах (на падали, в экскрементах, на высших грибах, пнях, в гнездах и т.д.) встречаются особые, часто очень богатые видами, но большей частью короткоживущие сообщества, на которые по-разному влияют различные биоценозы, в состав которых входят эти биохоры.
В каждую экосистему входят следующие функциональные компоненты:
а) продуценты – автотрофные организмы, которые с использованием солнечной энергии строят из неорганических соединений богатую энергией биомассу (зеленые растения, сине-зеленые водоросли, фото- и хемосинтезирующие бактерии);
б) консументы – гетеротрофные организмы, которые используют этот органический материал для получения и накопления энергии, изменяют или перестраивают органические вещества (животные и гетеротрофные микроорганизмы);
в) деструенты – гетеротрофы, которые разрушают использованные или отмершие остатки биомассы, разлагают их на неорганические составные части (минерализация), поступающие снова в резервуары минеральных веществ (бактерии, грибы).
Консументы питаются живым (биофаги) или мертвым (сапрофаги) органическим материалом. Среди биофагов могут быть выделены растительноядные организмы, или фитофаги (первичные консументы, к ним относятся также поражающие растения вирусы, бактерии, грибы и паразитические сосудистые растения), хищники (вторичные консументы, в их числе и паразиты первичных консументов) и конечные потребители – вершинные хищники (третичные консументы).
Представители рассмотренных выше трофических категорий (трофических уровней) связаны между собой односторонне направленной передачей биомассы в пищевые цепи (цепи питания). Все пищевые цепи входят в круговороты материи, ведущие от продуцентов к деструентам; цепи могут состоять из малого или большого числа звеньев.
Пастбищные пищевые цепи (рис. 12.14), или цепи хищников, начинаются с продуцентов: клевер – овца – волк; планктонные водоросли – дафния – плотва – щука – скопа. Для таких пищевых цепей характерно увеличение размеров особей (восходящие цепи) при одновременном уменьшении плотности популяций, скорости размножения и продуктивности по биомассе. Цепи паразитов могут начинаться с продуцентов (яблоня – щитовка – наездник) или консументов (овца – муха-кровососка – бактерии – вирусы); для них характерно повышение плотности популяции и скорости размножения при уменьшающихся размерах особей. Детритные цепи проходят через отмершую биомассу; они обычно коротки (листовой опад – дождевые черви – бактерии).
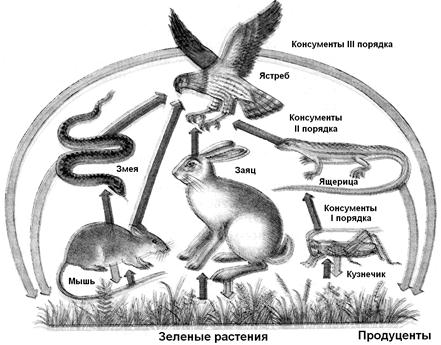
Рис. 12.14. Трофические цепи в наземном биогеоценозе
Так как продуцентов обычно поедают разные консументы, хищники используют много видов жертв, а всеядные организмы (человек, медведь, воробей) потребляют как продуцентов, так и консументов, т.е. живут на разных трофических уровнях, пищевые цепи многократно разветвляются и сплетаются в сложные пищевые сети.
Если расположить друг над другом графические отображения плотности популяций (число особей на 1 м2), биомассы (в граммах сухого или влажного веса на 1 м2) или продуктивности в энергетических эквивалентах (джоулях на 1 м2 в год) для всех членов каждого трофического уровня (в одной пищевой цепи), получится так называемая пищевая (экологическая) пирамида (рис. 12.15). Пирамиды энергии (так как в них учитывается время) стоят всегда «правильно». Напротив, «вверх ногами» стоят пирамиды чисел для цепей паразитов и цепей, начинающихся с древовидных продуцентов.

Рис. 12.15. Экологические пирамиды. А. Кузнечики поедают зеленые растения, лягушки – кузнечиков, змеи - лягушек, орел – змей. Внизу - пирамиды чисел (а), биомассы (б), энергии (в) для упрощенной модельной экосистемы: люцерна, телята, мальчик
12.5.2 Физиология экосистем
Экосистема - открытая система, в которой процессы образования и дальнейшей передачи биомассы идут за счет притока энергии солнечного излучения. При каждой передаче на следующий трофический уровень часть доступной энергии не воспринимается, часть отдается (в виде тепла),а часть расходуется на дыхание. По грубой оценке, при этом общая энергия каждый раз уменьшается в десять раз, а при передаче от первичных продуцентов консументам – даже в сто раз (рис. 12.4). Следствие этого – ограниченная длина пищевых цепей, а также уменьшение количества особей и общей энергии при переходе от низших уровней к высшим. Чем длиннее пищевая цепь, тем меньше остается к ее концу доступной энергии.
12.5.3 Развитие экосистем
Хотя экосистемы, находясь в стационарном состоянии, отличаютсястатистическим постоянством своих элементов, онимогут развиваться, переходя от более простых к более сложным формам.
Там, где отступающее море, высыхающее озеро, ледник или деятельность человека освобождают девственную сушу, а также там, где из вулканического пепла, из лавы или в результате выветривания горных пород образуется новая почва, начинается ее заселение редкой, одноярусной пионерной растительностью.С накоплением гумуса и пыли, с повышением влажности почвы развиваются двухъярусные вторичные сообщества, луга и степи. Устойчивая конечная стадия (климакс)этой первичной сукцессии – в большинстве случаев многоярусный лес (рис. 12.16). Вторичные сукцессии как бы залечивают повреждения, естественные или нанесенные человеком (последствия бури, вырубки, пожара, наводнения, выпаса скота). Каким будет конечный этап сукцессии, зависит прежде всего от макроклимата (климатический климакс). Например, на больших высотах и в высоких широтах климаксным сообществом будет уже не лес, а тундра. На сукцессию влияют также эдафические (почвенные, водные) и топографические особенности (уклон, экспозиция, уровень грунтовых вод). Так же может влиять деятельность человека – выпас скота, сенокос, полеводство (параклимакс), так что ландшафты чаще всего представляют собой мозаику из экосистем с разной степенью зрелости.
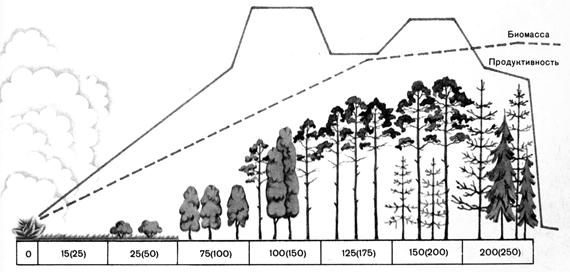
Рис 12.16. Схема сукцессии
В молодом, преимущественно автотрофном лесу производится излишек биомассы (Рп), который может накапливаться в виде древесины. Создающиеся ниши заселяются гетеротрофами, которые все больше используют прирост биомассы. Наконец, в климаксном лесу складывается сложная пищевая сеть с возрастающим удельным весом детритных пищевых цепей, в которых используется вся продукция. Поэтому человеку удается снимать высокие урожаи биомассы только на ранних стадиях сукцессии, когда нетто-продукция велика. В земледелии и лесном хозяйстве используются начальные фазы экосистем с немногими (предпочтительно одним)первичными продуцентами (монокультуры). Конечно, за это приходится расплачиваться неустойчивостью пионерных сообществ по отношению к внешним факторам и конкурирующим консументам. С гетеротрофами, желающими найти себе нишу в такой экосистеме, приходится бороться как с «вредителями».
Сукцессии экосистем, как и эволюция живого на всех уровнях, направлены на обеспечение дальнейшего существования, на гомеостаз. Очевидно, что стабильность подвижного равновесия при меняющихся условиях среды легче всего достигается в том случае, если экосистема состоит из максимально возможного числа компонентов (внутреннее разнообразие); тогда экологические потенции разных видов могут так дополнять друг друга, что разные помехи, как внешние (особенно непредсказуемые изменения абиотических факторов, к которым невозможно приспособиться), так и внутренние (чрезмерные репродуктивные потенциалы некоторых организмов), будут сглаживаться.
12.6 Человек и окружающая среда
Человек, будучи всеядным консументом, сначала занимал сравнительно скромное место в немногих стабильных экосистемах. Обладая выдающимся духовным потенциалом, изобретая и изготовляя орудия и оружие, он, однако, все более освобождался от своих естественных экологических связей и сам стал важнейшим экологическим фактором, из объекта эволюции превратился в ее субъект.Овладение огнем и развитие земледелия и скотоводства позволили ему отвлекать на себя энергетические потоки других экосистем. Так он научился использовать высокую продуктивность ранней фазы в сукцессиях естественных экосистем по возможности неограниченно в пространстве и времени. Для этого он стал сначала путем корчевания, выжигания, мелиорации или орошения сводить подходящие климаксные сообщества к начальным стадиям, а затем сделал эти стадии постоянными, поддерживая монокультуры, снимая почти весь урожай биомассы и ежегодно обновляя почвы вспашкой.
Более или менее параллельно с этими событиями человек узнал о фазе легкой запечатляемости стимул-объектов в ранней молодости у стадных животных. Эту особенность удалось использовать для их приручения, получив послушных илегко разводимых домашних животных. Промежутки между урожаями, определяемые сменой времен года, сделали необходимым появление экономики, основанной на запасах, и отсюда, видимо, возникла не оставляющая нас до сих пор вера в необходимость постоянного роста производства. Так была запущена та роковая спираль развития человечества, в которой производство излишков неизбежно ведет к дальнейшему экспоненциальному росту населения, а этот рост, в свою очередь, делает необходимым дальнейшее расширение производства.
Это стремление к максимизации и неестественно, и неэкологично, так как одностороннее развитие одного компонента со временем разрушает любую систему. Оно заставляет беспрерывно бороться с сопротивлениемсреды, которое вначале успешно противодействовало экспоненциальному росту человечества и его производства. Хотя в потенции каждый организм способен к экспоненциальному размножению, в природных экосистемах оно раньше или позже преобразуется под давлением противодействующих биотических и абиотических факторов (погодных явлений, врагов, паразитов, возбудителей болезней и т.д.) в логистический рост; в результате между всеми членами биоценоза устанавливается равновесие, в какой-то мере колеблющееся около средних значений и обеспечивающее стабильность биоценоза. Это неизбежный процесс, не требующий притока энергии извне.
Но если сукцессия искусственно останавливается на лабильной начальной стадии и при этом неуклонно повышается продукция постоянно изымаемой биомассы одного-единственного вида – культурного растения или домашнего животного, – приходится не только возмещать расходуемые минеральные вещества с помощью удобрений, но и вносить добавочную энергию, чтобы устранять противодействующие виды («сорняки», «вредители»), использующие монокультуру для себя, вынуждающие ее к логистическому росту или стремящиеся довести сукцессию до климакса. При этом в принципе безразлично, применяются ли против таких врагов химические (яды), биологические (другие, «полезные», организмы) или смешанные средства борьбы. В любом случае необходимо детальное знание биологии и экологических потенций всех компонентов системы, включая их способность к сопротивлению путем модификационной или мутационной адаптации.
Непосредственные враги самого человека – хищники и возбудители болезней (паразиты и патогены) – в результате охоты и достижений медицины (в том числе гигиены) настолько оттеснены, что хищники во всем мире практически уже не имеют для человека значения, а паразиты в высокоразвитых городских цивилизациях играют лишь подчиненную роль. Патогенные микроорганизмы – крошечные n -стратеги, обладающие огромным потенциалом размножения, который облегчает им возможность образования штаммов, резистентных к лекарственным веществам, и новых вирулентных штаммов, – все еще вызывают опасные эпидемии. И все же средняя продолжительность жизни человека выросла с 18 (в бронзовый век) до 70 лет (в Европе и Северной Америке). Это значительно способствовало демографическому взрыву XX века, в который, впрочем, особенно значительный вклад внесли развивающиеся страны Азии и Южной Америки. На повышение плотности населения (различное в разных регионах) накладывается все возрастающая урбанизация. Около 1800 г. в городах с населением 20 000 человек и больше жило менее 5% всего человечества, в 1950 г. – 21%, а сейчас – 30%. В начале ХІХ века на Земле было 27 крупных городов с населением 100 000 человек и больше, а в конце ХХ века – более 1800 таких городов.
До сих пор человек получал необходимую ему добавочную энергию почти исключительно из ископаемых энергетическихзапасов, накопившихся в форме угля, нефти и газа в результате образования биомассы в прошедшие эпохи. Но их исчерпание в обозримом будущем неизбежно, и потому необходимо искать другие источники энергии. Только благодаря ископаемым горючим материалам стали вообще возможными процессы индустриализации, которые косвенно способствовали развертыванию опасной спирали роста населения. В результате окружающей средой для человечества стала теперь практически вся биосфера, но для поддержания господства над ней человеку требуется все больше энергии. Хотя на Землю продолжают поступать огромные количества солнечной энергии, ни один технический процесс не смог пока удовлетворительно воспроизвести (а тем более превзойти) уникальную процедуру использования этой энергии в фотосинтезе зеленых растений. К тому же нам грозит и исчерпание невозобновляемых материальных ресурсов, например запасов урана, серебра, цинка.
Человек активно использует в своем сельском и лесном хозяйстве почти все наземные биомы. Но сейчас его деятельность затрагивает и практически все остальные естественные экосистемы – на них влияют хотя бы промышленные отходы, для которых в биосфере нет деструентов (например, пластмассы). Эти экосистемы уже изменены (ДДТ в организме антарктических пингвинов и во льду Арктики) или повреждены (загрязнение океанов нефтью, а атмосферы – SО2) антропогенными воздействиями и далеко отклонились от стационарного состояния. Поэтому все настойчивее выдвигается требование создать экономику, безвредную для природы, с ограниченным ростом при оптимальной численности населения. Но еще не известно, допускает ли вообще генетический потенциал человека такое качественное изменение жизненных потребностей, связанное с самоограничением и сокращением потребления, тем более что человек уже в такой значительной мере освободился от действия сил естественного отбора. До тех пор пока идеология роста производства даже охрану природы рассматривает лишь как средство к «лучшему использованию ресурсов» и стремится во всем мире не к оптимуму, а к максимуму, человечество не подойдет к «экологической морали», которая способствовала бы развитию экологической экономики, т.е. развитию цивилизации не за счет природы и против нее, а в устойчивой гармонии с нею.
Большие перспективы открывает контроль над рождаемостью, который уже сегодня привел в индустриальных обществах к стабилизации численности населения. С повышением уровня жизни в развивающихся странах такое положение может, наконец, стать всеобщей нормой. Но тогда постоянный рост имущества и власти, ресурсов и энергии перестанет быть мерой благосостояния. Ведь такое мировоззрение уже сделало человека единственным существом, наладившим организованное истребление себе подобных. Таким образом, всеобщую борьбу против войны можно рассматривать как многообещающий подход к человеческой экологии – к созданию самими людьми окружающей среды, более достойной человека.
Л1(17). ИСОН с высоким уров
Дата публикования: 2014-10-19; Прочитано: 2146 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
