 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Мембранный потенциал
|
|
Диффузионный потенциал на границе двух растворов постепенно уменьшается в результате выравнивания концентраций. Стабилизировать потенциал, возникающий на границе раздела жидкость - жидкость, можно, если соприкасающиеся растворы разделить мембраной с избирательной проницаемостью (полупроницаемой). Такая мембрана способна избирательно пропускать те или иные ионы и молекулы, в результате чего возникает мембранный потенциал - фм.
Мембранным потенциалом называется потенциал, возпикающий между сторонами мембраны с избирательной проницаемостью, разделяющей два раствора различного co става.
 |
Величину мембранного потенциала можно определить, составив гальваническую цепь, в которой в растворы, разделенные мембраной с избирательной проницаемостью, опущены два электрода сравнения:
ЭДС такой гальванической цепи характеризует величину мембранного потенциала.
Мембранный потенциал зависит от отношения активностей ионов в растворах, разделенных мембраной, и от свойств мембраны. Мембраны характеризуются проницаемостью, т. е. способностью пропускать определенные виды ионов, которые являются потенциалопределяющими в возникновении данного мембранного потенциала. Проницаемость мембраны для разных ионов X и Y характеризуется коэффициентами проницаемости Р(Х) и P(Y). Рассчитать значение мембранного потенциала для мембраны, проницаемой для двух видов ионов, можно по следующему уравнению:
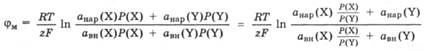
где анар(Х), авн(Х), aнap(Y), aBH(Y) - активность потенциал определяющих ионов X и Y в растворах снаружи и внутри клетки; Р(Х), P(Y) -коэффициент проницаемости мембраны для ионов X и Y.
Для живых клеток, особенно для клеток нервной системы, важное значение имеет различие в концентрациях ионов К+ и Na+ внутри и снаружи клетки, поэтому эти ионы являются птенциалопределяющими для клеток нервной системы:
| Ионы | авн(Х), ммоль/л | aнар(X), ммоль/л | авн(Х)/анар(Х) | P(K+)/P(Na+) |
| покой | возбуждение | |||
| к+ Na+ | 400 50 | 20 440 | 20 1/9 | 100 1/12 |
Через клеточную мембрану ионы К+ и Na+ самопроизвольно передвигаются по ионным каналам в соответствии с градиентом концентраций. В состоянии покоя в мембране в основном открыты каналы для прохождения ионов К+ и практически закрыты натриевые каналы. При возбуждении - наоборот: открыты главным образом каналы для ионов Na+ и почти полностью закрыты для ионов К+. Таким образом, проницаемость клеточной мембраны для ионов К+ и Na+ зависит от ее состояния: покой или возбуждение - и характеризуется различным отношением коэффициентов проницаемости для этих ионов P(K+)/P(Na+).
Избирательная проницаемость клеточной мембраны и разница в активности ионов Na+ и К+ по обе стороны от нее приводят к установлению мембранного потенциала (рис. 25.6).
В покое ионы К+ из внутриклеточного раствора, где их концентрация в 20 раз выше, чем снаружи, переходят через клеточную мембрану в наружный раствор. При этом наружная поверхность мембраны заряжается положительно за счет перешедших ионов К+, а внутренняя поверхность - отрицательно за счет избытка органических анионов, оставшихся внутри клетки. Таким образом, возникает мембранный потенциал покоя фпок, препятствующий дальнейшему выходу ионов К+ из внутриклеточного в наружный раствор и установлению равновесия в их движении.
Потенциалом покоя называется мембранный потенциал, возникающий между внутренней и наружной сто- I ронами клеточной мембраны, находящейся в невозбужденном состоянии.
Определить потенциал покоя можно с помощью двух микроэлектродов сравнения, вводимых внутрь клетки и в наружный раствор. Измеренное значение фпок для различных клеток лежит в пределах от -70 до -90 мВ. Знак минус говорит о том, что внутренняя поверхность мембраны заряжежа отрицательно. Учитывая активности ионов Na+ и К+ внутри клетки и снаружи, а также отношение коэффициентов проницаемости мембраны для этих ионов, вычислим потенциал покоя:
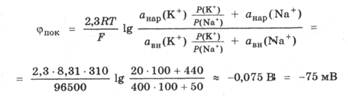
Расчетное значение фпок хорошо согласуется с: экспериментально измеряемым.
При раздражении клетки химическим, электрическим или механическим воздействием она переходит в возбужденное состояние, при этом проницаемость ее мембраны для ионов Na+ становится значительно выше, чем для К+. Поэтому ионы Na+ из наружного раствора, где их концентрация в 9 раз выше, чем внутри клетки, устремляются через клеточную мембрану во внутренний раствор. Ионы Na+ переносят положительный заряд с наружной поверхности мембраны и перезаряжают ее внутреннюю поверхность, меняя знак заряда с "-" на "+" и вызывая быструю деполяризацию мембраны (рис. 25.6). При этом на короткое время (< 10-3 с) мембранный потенциал становится равным примерно +40 - +60 мВ, что полностью согласуется с экспериментальными и расчетными данными. После прекращения возбуждения мембрана вновь становится проницаемой для ионов К+ и непроницаемой для Na+. Ионы К+ опять выходят из клетки в соответствии с градиентом концентрации, унося с собой положительный заряд до тех пор, пока на мембране не восстановится потенциал покоя, т. е. не произойдет реполяризация мембраны.
Таким образом, при возбуждении клеточной мембраны за (1-2) • 10-3 с ее потенциал с отрицательного значения (= -80 мВ)
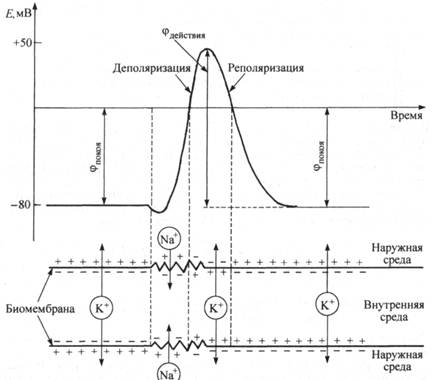
Рис. 25.6. Возникновение мембранных потенциалов покоя и действия
меняется на положительный (< +50 мВ), а затем вновь возвращается к первоначальному значению.
Потенциалом действия называется амплитуда колебания (деполяризация и реполяризация) мембранного потенциала, возникающая при возбуждении клеточной мембраны.
На рис. 25.6 схематично показано возникновение потенциала действия в клетке. Амплитуда колебания потенциала составляет примерно 120-140 мВ.
Потенциал действия, возникнув на одном участке клетки, вызывает возбуждение соседних участков и распространяется по всей поверхности мембраны со скоростью 1-110 м/с.
Количество ионов К+ и Na+, проходящих через мембрану во время генерации потенциала действия, составляет не более чем 1 • 10-7 от количества этих ионов внутри клетки, но даже и при прохождении большого числа импульсов концентрации ионов К+ и Na+ в растворах по обе стороны мембраны остаются практически постоянными. Это происходит потому, что в мембране клетки действует Na+/K+-нacoc. Он, используя энергию АТФ, выкачивает из клетки ионы Na+ и накачивает в нее ионы К+ (в соотношении 3: 2) против градиента концентраций этих ионов (разд. 13.1.1).
Следует отметить, что в общем случае возникновение потенциала покоя и генерация потенциала действия на мембранах различных клеток связаны с переходом через мембрану не только ионов К+ и Na+, но также Са2+, С1-, Н+ и других ионов.
Изучение биомембран мышечных и секреторных клеток показало, что у многих из них потенциал покоя возникает за счет перемещения ионов Na+, а потенциал действия имеет кальциевую природу. В этом случае генерация потенциала действия происходит при возбуждении в результате открывания кальциевых каналов и перемещения ионов Са2+ внутрь клетки, что приводит к сокращению мышцы или к выбросу секрета.
Современные исследования биологических внутриклеточных мембран показали, что на них возникает протонный потенциал из-за различия в концентрациях ионов водорода в растворах, разделенных этими мембранами (разд. 9.3.4). Протонный потенциал при условии проницаемости внутриклеточной мембраны только для ионов Н+ можно вычислить по уравнению для расчета мембранного потенциала, введя в него водородный показатель рН = -lg а(Н+).
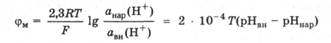
Установлено, что протонный потенциал может служить источником энергии для всех видов работ, характерных для живой системы: химической, осмотической, механической, - и источником теплоты.
Следует отметить, что работа клеток нарушается, если изменяется ионная проницаемость клеточных мембран. Подобное может происходить, например, под действием некоторых ядов: в нервных клетках при возбуждении блокируются каналы для прохождения ионов Na+, и поэтому прекращается генерация и передача потенциала действия вдоль нервного волокна. Этим и объясняется токсическое действие ядов на нервную систему организма (разд. 11.4).
Генерирование мембранного потенциала связано с работой сердца, мозга, мышц. Электрические потенциалы, возникающие при деятельности сердца, можно регистрировать с помощью электрокардиографа на электрокардиограмме. ЭКГ - важнейшая характеристика сердечной деятельности. Биоэлектрические потенциалы мозга регистрируются на электроэнцефалограмме, мышц - на электромиограмме, желудка - на электрогастрограмме и т. д.
Избирательная проницаемость мембран относительно определенного вида ионов и зависимость мембранного потенциала от концентрации этих потениалопределяющих ионов лежат в основе работы ионоселективных электродов. Эти электроды позволяют измерять концентрацию данного потенциалопределяющего иона в исследуемой системе по величине возникающего мембранного потенциала на электроде. Ионоселективные электроды широко используются в потенциометрии.
Дата публикования: 2014-10-16; Прочитано: 4618 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
