 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Первичная структура нуклеиновых кислот. Нуклеиновые кислоты – высокополимерные соединения, состоящие из очень большого числа отдельных мононуклеотидов
|
|
Нуклеиновые кислоты – высокополимерные соединения, состоящие из очень большого числа отдельных мононуклеотидов, т.е. представляют собой полинуклеотиды. По химической структуре нуклеиновые кислоты состоят из цепочек нуклеотидов — чередующихся остатков пентозы — рибозы или дезоксирибозы и фосфорной кислоты (фосфата) с азотистыми основаниями, присоединенными к углеводным фрагментам N-гликозидной связью.
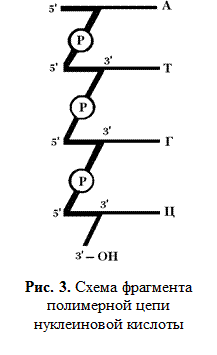
Мононуклеотиды связываются в нуклеиновые кислоты путем образования сложноэфирных связей между 3'-углеродным атомом пентозы одного мононуклеотида, фосфатом и 5'-углеродным атомом пентозы другого нуклеотида (рис. 2). В молекуле рибонуклеиновой кислоты (РНК), таким образом, связываются третий и пятый атомы углеродов рибозы, а в дезоксирибонуклеиновой кислоте (ДНК) — пятый и третий атомы углерода дезоксирибозы.
 Разные нуклеиновые кислоты отличаются друг от друга числом азотистых оснований в цепи.
Разные нуклеиновые кислоты отличаются друг от друга числом азотистых оснований в цепи.
Для обозначения первичной структуры нуклеиновых кислот пользуются однобуквенными символами нуклеозидов (русскими или латинскими): А (А) - аденозин, Г (G) - гуанозин, Ц (С) - цитидин У (U) -уридин, Т (Т) - тимидин.
Из четырех разных нуклеотидов может быть построено огромное количество вариантов нуклеиновых кислот.
Конец нуклеотидной цепи, у которой остается свободной гидроксильная ОН-группа, у третьего углеродного атома рибозы называется 3'-концом, а другой конец цепи, у которого пятый углеродный атом рибозы соединен с остатком фосфорной кислоты, называют 5'-концом.
Таким образом, первичная структура нуклеиновых кислот — это последовательность расположения нуклеозидфосфатов в линейной цепи, на одном конце которой находится 5'-фосфатная группа (это 5'-конец), а на другом — свободная 3'-ОН-группа (3'-конец).
На схеме (рис. 3) показаны связи между отдельными мононуклеотидами в молекулах РНК и ДНК.
Специфичность нуклеиновых кислот определяется не только их нуклеотидным составом, но и последовательностью отдельных нуклеотидов в цепи нуклеиновых кислот. ДНК каждого определённого вида характеризуется только ей присущей специфичной последовательностью нуклеотидов, и это определяет биосинтез специфичных для каждого вида организма молекул белков. Состав ДНК совершенно не зависит от условий среды, в которой выращиваются растения, он остается строго постоянным даже при резко различных условиях их питания
 Вторичная структура нуклеиновых кислот
Вторичная структура нуклеиновых кислот
Вторичная структура ДНК. Состав и содержание нуклеотидов в молекулах ДНК изучены очень детально. Исследования показали, что содержание отдельных оснований в ДНК различных организмов подчиняется строгим закономерностям. Особенность нуклеотидного состава ДНК выражена в правилах Чаргоффа:
1. Сумма пуриновых оснований (Пур) равна сумме пиримидиновых оснований (Пир), т. е. А+Г = Т+Ц,илиПур = Пир, или 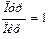 ;
;
2. Содержание аденина равно содержанию тимина, а содержание гуанина равно содержанию цитозина, т.е., А = Т, или 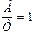 , а Г = Ц, или
, а Г = Ц, или 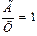 .
.
В отличие от этого количество пар оснований Г+Ц и А+Т может быть различным.
В зависимости от преобладания той или иной пары оснований различают АТ-тип или ГЦ-тип ДНК. У ДНК АТ-типа А+Т > Г+Ц, т. е. сумма аденина и тимина превышает сумму гуанина и цитозина, а у ДНК ГЦ-типа А+Т < Г+Ц, т. е. сумма аденина и тимина меньше суммы гуанина и цитозина.
Отношение  одна из важных характеристик ДНК и показатель ее специфичности по нуклеотидному составу. Каждый вид организмов имеет свойственный для данного вида нуклеотидный состав ДНК, причем у близких видов различия в составе ДНК обычно значительно меньше, чем у далеко отстоящих в систематическом отношении.
одна из важных характеристик ДНК и показатель ее специфичности по нуклеотидному составу. Каждый вид организмов имеет свойственный для данного вида нуклеотидный состав ДНК, причем у близких видов различия в составе ДНК обычно значительно меньше, чем у далеко отстоящих в систематическом отношении.
Основываясь на этой закономерности, не свойственной молекулам РНК, и данных рентгеноструктурных анализов, Дж. Уотсон и Ф. Крик в 1953 г. предложили модель строения вторичной структуры ДНК в виде двух правозакрученных спиральных полинуклеотидных цепей, переплетенных друг с другом и противоположно направленных. Эта структура двойной спирали стабилизирована межцепочечными водородными связями между специфическими парами азотистых оснований, которые направлены от сахарофосфатного остова каждой цепи внутрь спирали перпендикулярно ее оси.
Аденин всегда связан двумя водородными связями с тимином, а гуанин — тремя водородными связями с цитозином (рис. 4). Это единственно возможное соединение нуклеотидов в двойной цепи, так как два пуриновых основания не могут совместиться в
 |
пространстве, а два пиримидиновых основания будут расположены слишком далеко, чтобы между ними могли образоваться водородные связи. Эти пары называют комплементарными парами оснований, благодаря которым первичная структура одной цепи ДНК комплементарна первичной структуре другой цепи. Две цепи молекулы ДНК дополняют друг друга, и порядок оснований в одной цепи позволяет совершенно точно предсказать порядок их расположения в другой. Например, если в одной цепи имеется последовательность АТГТЦ, то в другой возможна только последовательность ТАЦАГ.
Плоскости пиримидиновых оснований перпендикулярны оси спирали и параллельны друг другу, образуя стопку оснований. Между стопками возникают гидрофобные взаимодействия, вносящие основной вклад в стабилизацию двойной спирали, больший, чем водородные связи между цепями. Пентозофосфатные части цепей располагаются по периферии спирали, образуя ее ковалентный остов (рис. 5).
Двойная спираль ДНК существует в разных формах, обозначенных на рис. 5, в: 1...3.
В-форма ДНК имеет диаметр спирали 2,1 нм, 10 пар оснований на виток, шаг спирали 3,4 нм; пары оснований перпендикулярны продольной оси спирали; продольная ось спирали пронизывает стопку оснований посередине. Между цепями расположены две «бороздки» — большая и малая (рис. 6).
А-форма ДНК имеет диаметр спирали 2,5 нм, 11 пар оснований на виток, шаг спирали 2,8 нм; пары оснований наклонены под углом 20° к плоскости, перпендикулярной продольной оси спирали; они обвивают продольную ось так, что внутри нее вдоль имеется полость диаметром 0,8 нм. Также есть две «бороздки», но большая шире и глубже, чем в В-форме.
 |
С-форма ДНК имеет 9,3 пары оснований на виток, которые наклонены под углом 6° к плоскости, перпендикулярной оси спирали, и немного — относительно друг друга.
Z-форма ДНК в отличие от других имеет левую спираль с 12 парами оснований на виток, а пентозофосфатные цепи имеют не плавные линии, как в В- или С-формах, а ломаный Z-образный вид.
Форма спирали ДНК, по-видимому, зависит от степени гидратации молекулы: А-форма (кристаллическая) существует при массовой доле воды не выше 40 %; В-форма (паракристаллическая) — при массовой доле воды более 40%. Предполагается, что в метаболически активных тканях преобладает В-форма.
Основная часть ДНК в клетке сосредоточена в ядрах (обычно около 30% на сухую массу). В ядрах она связана с белками основного характера — гистонами. ДНК присутствует также в хлоропластах зеленых растений. Некоторое ее количество найдено в митохондриях.
 Содержание ДНК в клетке всегда постоянно и зависит лишь от числа хромосом. Молекулярная масса ДНК, выделенной из различных объектов, достигает 108—109 и даже больше. Её молекулы построены из сотен тысяч и даже миллионов пар нуклеотидов.
Содержание ДНК в клетке всегда постоянно и зависит лишь от числа хромосом. Молекулярная масса ДНК, выделенной из различных объектов, достигает 108—109 и даже больше. Её молекулы построены из сотен тысяч и даже миллионов пар нуклеотидов.
Вторичная структура РНК. Рибонуклеиновые кислоты, так же как и ДНК, – полимерные соединения, состоящие из рибонуклеозидмонофосфатов (рибонуклеотидов). В клетках любых биологических объектов содержится три основных типа РНК: рибосомная РНК (рРНК), транспортная РНК (тРНК) и информационная, или матричная, РНК (мРНК). Все три типа РНК различаются по свойствам, строению, функциям и локализации в клетках (табл. 1). В большинстве клеток содержание РНК в 5—10 раз превышает содержание ДНК.
На рибосомную РНК приходится основная часть РНК клетки (до 80—90%). Она содержится в рибосомах — внутриклеточных частицах, принимающих участие в биосинтезе белка. По сравнению с другими типами имеет наибольшую молекулярную массу и характеризуется метаболической устойчивостью. В рибосомах высших организмов содержится четыре вида рибонуклеиновой кислоты, два вида высокомолекулярной рРНК и два вида низкомолекулярной рРНК. Эти четыре вида рРНК содержатся в двух субъединицах рибосом. Они имеют разное соотношение нуклеотидов и различаются по последовательности оснований в цепях.
Таблица–1 Некоторые характеристики РНК
| Показатель | мРНК(иРНК) | тРНК | рРНК |
| Массовая доля в клетке, % от суммы | ~2 | 10...15 | 80...90 |
| Число нуклеотидов в молекуле | 75...3000 | 70...90 | 1500...4500 |
| Молекулярная масса, Да | 25·10-3...1·10-6 | 2,5·104...3,1·104 | 0,3·106...1,6·106 |
| Форма молекулы | Вытянутая | «Клеверный лист» | Компактная |
| Коэффициент седиментации | (6...25)S | 4S | 28S, 18S, 16S |
Транспортная РНК составляет обычно 10—15% всей РНК клетки, имеет сравнительно низкую молекулярную массу. Она не связана с клеточными структурами и находится в клетках в растворенном состоянии. Функция транспортной РНК заключается в переносе, транспорте аминокислот к месту синтеза белка. Для переноса каждой аминокислоты существует своя собственная тРНК, а для некоторых из них — по нескольку тРНК, и общее число видов тРНК доходит до 60.
В составе рРНК и тРНК кроме нуклеотидов (Г, А, Ц и У) содержатся и минорные нуклеотиды, наибольшее количество которых обнаружено в тРНК. Некоторые минорные нуклеотиды, содержащиеся в малых количествах по сравнению с главными (псевдоуридин), содержат метилированные или дигидропроизводные главных оснований.
Матричные РНК составляют около 2 – 5% всей РНК клетки. В отличие от других типов РНК она содержит только четыре основания — А, Г, Ц, У. Нуклеотидный состав ее подобен составу одного из участков цепи ДНК. Иногда мРНК называют иРНК (информационные РНК). Функции мРНК заключаются в переносе информации от ДНК на синтезируемый белок. В клетках синтезируются тысячи различных белков, и состав каждого из них определяется специфической мРНК или специфическим участком ее полинуклеотидной цепи. Таким образом, существует огромное количество мРНК, отличающихся друг от друга первичной структурой – по числу разных белков в организме, и общее число видов мРНК очень велико.
Молекулы РНК построены из одной полинуклеотидной цепи, но отдельные их участки могут образовывать двойные спирали из-за присутствия комплементарных нуклеотидных пар А—У и Г—Ц. Такие спирализованные участки называют шпильками, включающими 20...30 нуклеотидных пар. Они чередуются с неспирализованными участками. Транспортная РНК имеет вторичную структуру, которая представляет собой четыре спирализованных участка и три или четыре одноцепочечные петли — так называемый «клеверный лист» (рис. 7). На ряде участков молекулы между соответствующими парами оснований образуются водородные связи, которые обусловливают определенную конфигурацию тРНК. Кроме этого, полинуклеотидная цепь молекулы, сворачиваясь на себя, образует двойные спирали. Следует обратить внимание на тройку оснований, которая находится в нижней петле молекулы тРНК. Эти тройки оснований, которые называют антикодонами, индивидуальны для каждого вида тРНК, они определяют биосинтез специфических белков, характеризующихся строго определенной последовательностью аминокислот.
Дата публикования: 2015-11-01; Прочитано: 1309 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
