 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Неорганических соединений азота
|
|
Для построения органических веществ нужны в первую очередь кислород, водород, углерод, азот, сера, фосфор и некоторые другие химические элементы. Растения не испытывают недостатка в кислороде и водороде, получая их из воздуха и воды. Запасы азота в природе тоже неисчерпаемы, ведь 4/5 объема окружающего нас воздуха состоит из этого элемента. Столб воздуха над гектаром почвы содержит до 80 тысяч тонн азота. Весьма значительны запасы азота и в почве, а между тем именно в нем органическая жизнь чаще всего испытывает острую нужду. И плохая урожайность сельскохозяйственных культур связана, в первую очередь, с недостатком азотистого питания. Это происходит потому, что растениями может быть усвоена только небольшая группа азотистых соединений. И свободный азот атмосферы, и многие формы связанного азота не могут служить для них питанием. Только специальные физиологические группы микроорганизмов обладают способностью фиксировать и трансформировать азот, переводить его в минеральную форму, после чего он становится доступным для растений. В зеленых частях растений происходит синтез органических азотистых соединений, которые усваивают соли азотной кислоты, содержащиеся в почве. Животные питаются органическими веществами, образованными растениями, и используют их для синтеза своих белковых веществ.
Цикл трансформации азотсодержащих веществ в природе состоит из четырех взаимосвязанных процессов: биологической фиксации атмосферного азота, аммонификации, нитрификации и денитрификации.
Биологическая фиксация молекулярного азота. Биологическая фиксация молекулярного азота атмосферы, который находится в недоступной для высших растений форме, является одним из главнейших источников пополнения запасов азота в почве. Биологической фиксацией молекулярного азота обладают микроорганизмы, получившие название азотфиксирующих. Они фиксируют азот при обычном атмосферном давлении и нормальной температуре. В промышленных условиях фиксация азота идет химическим путем при температуре 400°С-500°С и давлении 35 МПа, что требует затраты большого количества энергии. Процесс биологической фиксации азота удовлетворяет около 20% потребности растений. По приблизительным подсчетам, на пашей планете почвенные микроорганизмы фиксируют ежегодно до 130 млн. тонн, а водные микроорганизмы - до 190 млн. тонн азота, в то время как мировое производство азотных удобрений пока не превышает 60-70 млн. тонн. Следует при этом отметить, что микробиологическая азотфиксация является источником дешевого, безвредного для окружающей среды, биологического азота.
Азотфиксирующие свободно живущие микроорганизмы ежегодно переводят в почву по 15-18 кг азота на 1 га, а микроорганизмы, живущие в симбиозе с растениями (клубеньковые бактерии), накапливают в почве до 70-80 кг азота на 1 га.
Типичными представителями свободноживущих азотфиксирующих микроорганизмов являются бактерии родов Clostridium и Azotobacter. Большую роль в биологической азотфиксации играют сине-зеленые водоросли.
Суть химизма биологической азотфиксации состоит в постепенном разрыве тройной связи, которой связаны атомы в молекуле азота. Наиболее трудно разрывается первая связь, намного легче разрываются вторая и третья. Постепенный разрыв этих связей осуществляется нитрогеназной мультиферментной системой азотфиксирующих микроорганизмов. При этом освободившийся атом азота замещается атомом водорода. Поэтому фиксация азота представляет собою восстановительный процесс. На восстановление одной молекулы азота до аммиака затрачивается 12 молекул АТФ. Энергию, затрачиваемую на этот процесс, свободно живущие азот-фиксаторы добывают за счет реакции катаболизма (от греч. katabole - распад), т.е. распада сложных органических соединений.
Конечным продуктом восстановления молекулы азота является аммиак, который накапливается в окружающей среде и подвергается нитрификации. В ходе этого процесса образуются промежуточные продукты, которые не накапливаются в окружающей среде, а преобразуются в конечный продукт (аммиак) на одном и том же ферментном комплексе:
N  N
N  HN = NH H2N - HN2 2NH3
HN = NH H2N - HN2 2NH3
Азот Диамид Гидразин Аммиак
Симбиотические азотфиксирующие микроорганизмы представлены клубеньковыми бактериями, которые тесно связаны с миром высших растений. Они выполняют грандиозную по своим масштабам работу разрушения и минерализации органических веществ.
В процессе роста растений на поверхности корней и надземных частей выделяются органические вещества, синтезированные растительным организмом. Образуется так называемый экзосмос растений, содержащий органические кислоты, витамины, ростовые вещества и т.д. Это создает благоприятные условия для жизнедеятельности на корнях и надземных частях растений обильной сапрофитной микрофлоры. Эта микрофлора перерабатывает органические и минеральные соединения, делая их доступными для растений, синтезирует ростовые вещества (гибберелин и гетероауксин), вырабатывает антибиотические вещества, подавляющие рост фито патоген ной микрофлоры. Поэтому корневая система растений всегда обильно заселена микроорганизмами. Они располагаются как на поверхности корней, так и в почве, прилегающей к корням. Эта область носит название ризосфера (греч. rtiiza - корень), а клубеньковые бактерии объединены в обширный род Rhisobium.
Особенно обильно обогащают почву азотом бобовые растения. Это явление было отмечено еще древнеримским философом Варроном за 37 лет до н.э. В 1838 году, французский ученый Ж.Б. Буссенго установил, что содержание азота в бобовых растениях значительно превосходит количество, которое они могли получить из почвы и воды. И только в 1866 году, русский ученый М.С. Воронин описал бактерии, обнаруженные им в клубеньках бобовых растений, и высказал предположение об их непосредственной связи с образованием клубеньков. Выделить бактерии из клубеньков удалось в 1888 году. Бейеринку и С.Н. Виноградскому.
В дальнейшем была установлена способность микробов в симбиозе с бобовыми фиксировать молекулярный азот из воздуха. Это было великое открытие. Механизм процесса азотфиксации клубеньковыми бактериями подробно изучен японским ученым Кубо. Он обнаружил, что клубеньки имеют розовый цвет; и установил, что он обусловлен пигментом леггемоглобином. Он назвал его гемоглобином клубеньковых растений. Этот пигмент образуется только при симбиозе бобовых растений и клубеньковых бактерий, что позволяет им фиксировать в среднем до 200 кг азота на 1 га, причем 2/3 его они берут из воздуха, а 1/3 из почвы. При этом клубеньковые бактерии используют соединения углерода, синтезированные бобовыми растениями, энергию АТФ из митохондрий клетки-хозяина.
Однако, несмотря на высокую эффективность фиксации азота, симбиотические азотфиксирующие микроорганизмы в общем азотном балансе биосферы занимают лишь второе место, уступая первое место свободноживущим азотфиксаторам. Это определяется, прежде всего, незначительной долей бобовых растений в естественных биоценозах. Нередко в экосистемах бобовые вовсе отсутствуют.
Формировать клубеньки и фиксировать азот воздуха в симбиозе с микроорганизмами могут не только бобовые растения. На корнях ольхи, березы, облепихи, хвойных и других растений имеются образования, подобные клубенькам бобовых, симбионтами в которых являются микроскопические грибы. По способности фиксации азота такие растения не уступают бобовым.
После выделения Бейеринком чистой культуры клубеньковых бактерий родилась идея использовать их для улучшения образования клубеньков, усиления фиксации атмосферного азота, а следовательно, повышения урожайности. С этой целью в 1896 году в Германии получены препараты клубеньковых бактерий под названием «нитрагин». В настоящее время получение препаратов клубеньковых бактерий поставлено на промышленную основу.
Аммонификация. Разнообразные органические азотсодержащие соединения составляют до 99% всего запаса азота в почве. Первое место среди них занимают белки, на долю которых приходится не менее 50% от органических соединений.
Превращение органического азота в неорганический (аммиачный) происходит при гниении белков. Попадая в почву, белковые вещества становятся субстратом для обильного размножения в них гнилостных микробов, которые посредством своих протеолитических экзоферментов расщепляют молекулу белка на более простые соединения - пептиды и олигопептиды. Эти соединения проникают в бактериальную клетку через цитоплазматическую мембрану, где расщепляются эндоферментами пептидазами до аминокислот. Разрушение аминокислот в аэробных условиях начинается с их дезаминирования, т.е. отщепления аминогруппы от молекулы аминокислоты, что приводит к выделению аммиака:
R-CHNH2-COOH + О2 R-COOH + СО2 + NH3
В аэробных условиях процесс дезаминирования аминокислот идет энергично с образованием аммиака, углекислого газа, воды и сульфатов. В анаэробных условиях некоторые аминокислоты подвергаются декарбоксилированию с образованием углекислоты и первичных аминов, к числу которых относятся такие высокотоксичные соединения, как кадаверин, путресцин и агматин, известные под названием трупных ядов. Анаэробное разрушение аминокислот сопровождается накоплением в окружающей среде специфических веществ с резким неприятным запахом - запахом аммиака, индола, скатола, меркаптана, летучих жирных кислот и другие. Конечными продуктами анаэробного распада являются аммиак, сероводород, углекислый газ, водород и вода.
У аммонифицирующих бактерий имеется своя специализация, то есть одни микробы разлагают белок до аминокислот, другие продолжают это расщепление до более простых соединений, включаясь на разных стадиях этого процесса. В аэробных условиях аммонификацию белков начинают неспороносные бактерии из родов Micrococcus, Pseudomonas, Proteus, а также микобактерии и плесневые грибы. На смену им приходят различные виды бацилл (Bac.subtiiis, Bac.megaterium, Вас. mesentericus, Bac.mycoides и др.). На завершающих этапах, аэробной аммонификации подключаются актиномицеты. В анаэробных условиях процесс аммонификации осуществляется клостридиями.
Бактериальному дезаминированию подвергаются органические азот- содержащие вещества небелковой природы, такие, как мочевина, мочевая и гиппуровая кислоты, входящие в состав мочи человека и животных. Взрослый человек выделяет ежедневно до 50 г мочевины. Если учесть все население земного шара и животный мир, то ежедневно на планете образуются миллионы тонн мочевины, не пригодной для питания растений и животных. Без микроорганизмов это колоссальное количество связанного азота было бы изъято из биологического цикла. Этого не происходит благодаря разложению мочевины, главным образом, за счет деятельности специфичных аэробных бактерий. Под действием фермента уреазы, выделяемой микроорганизмами, наблюдается гидролиз мочевины с образованием углекислого аммония, который почти тотчас же разлагается на составные компоненты: аммиак, углекислоту и воду:
Н2О
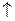
CO(NH2)2 + 2Н2О (NH4)2CO3 CO2

2NH3
Бактерии, разлагающие мочевину, получили название уробактерий. Специфической особенностью уробактерий является их способность развиваться в щелочной среде при значении рН 9-10 (алкилофильные бактерии).
Таким образом, в процессе трансформации азота за счет аммонификации белков и других азотсодержащих органических соединений небелковой природы образуется аммиак. Благодаря аммонифицирующим микробам от умерших организмов очищается поверхность земли, вода рек, озер, морей и других водоемов. Образовавшиеся в результате гниения углекислый газ, аммиак и вода используются для построения новой органической жизни.
Нитрификация. Процесс нитрификации - второе важное звено в цикле трансформации азотсодержащих веществ. Аммиак, образующийся в процессе аммонификации в почве и воде, сравнительно быстро окисляется нитрифицирующими бактериями до нитритов и нитратов. Нитраты являются одной из форм минерального азота, необходимого для питания растений. Накопление нитритов и нитратов в почве вызывает подкисленне почвенного раствора, что положительно сказывается на повышении растворимости некоторых соединений фосфора и железа и способствует лучшему усвоению их растениями.
О том, что в процессе нитрификации участвуют бактерии, впервые отмечено Л. Пастером. Позднее, в 1890 - 1892 гг., С.Н.. Виноградский выделил чистую культуру нитрифицирующих бактерий и установил, что процесс нитрификации протекает в два этапа. На первом этапе аммиак окисляется до азотистой кислоты по следующему уравнению:
2NH3 + ЗО2 = 2НNО2 + 2Н2О + 663,6 кДж.
В этом процессе участвуют нитритные (или нитрозные) бактерии, относящиеся к родам Nitrosomanas, Nitrosococcus, Nitrosolobus. Nitrosospira. Они разлагают аммиак до гидрокс ил амина, который затем окисляется до нитрита. Энергия, высвобожденная в реакциях окисления, аккумулируется в АТФ и в дальнейшем используется для фиксации углекислоты и других биосинтетических процессов.
NH3 + О2 + HAD  H2
H2  NH2OH + Н2О + HAD+
NH2OH + Н2О + HAD+
гидроксиламин
NH2OH + О2 NO2 + Н2О + Н+
NH2OH HNO + 2е- + 2Н+
нитроксил 
в АТФ
На втором этапе нитрификации азотистая кислота окисляется в азотную:
2HNO2 + O2= 2HNO3 + 201,6 кДж.
Второй этап нитрификации - окисление нитритов в нитраты -осуществляют нитратные бактерии родов Nitrobacter, Nitrospina, Nitrococcus. Все нитрифицирующие бактерии - облигатные аэробы.
Денитрификация. Нитраты в почве расходуются весьма интенсивно. Часть их потребляется в процессе питания растений, часть вымывается из почвы, а некоторое количество нитратов усваивается почвенными микроорганизмами при синтезе белка цитоплазмы. Многие почвенные микроорганизмы способны восстанавливать соли азотной кислоты (нитраты) до свободного азота. Этот процесс называется денитрификацией:
NaNO3 NaNO2 NH3 N2
Денитрифицирующие бактерии используют нитраты в качестве акцептора водорода при отсутствии молекулярного кислорода для окисления органических веществ до углекислого газа и воды и для получения энергии.
Восстановление нитратов и нитритов до газообразного азота происходит в результате прямой и косвенной денитрификации. Прямая или биологическая денитрификация наиболее распространена в природе и связана с жизнедеятельностью факультативных анаэробов, которые при недостатке молекулярного кислорода используют кислород нитратов и нитритов для энергетических процессов, а азот этих соединений - для синтеза белка.
Прямая денитрификация подразделяется на ассимилляционную и диссимилляционную. При ассимиляционной денитрификации нитраты потребляются в качестве источника азота и восстанавливаются до аммиака, который расходуется клеткой в процессе биосинтеза. В процессе диссимиляционной денитрификации нитраты служат акцептором водорода, т.е. восстанавливаются и используются в качестве источника кислорода. Конечными продуктами восстановления нитратов являются молекулярный азот, оксид азота (I) или оксид азота (II). В ходе этого процесса выделяется энергия, используемая микроорганизмами. Поэтому денитрификацию называют иногда процессом нитратного дыхания микроорганизмов. Способностью диссимиляционной денитрификации обладают мезофильные аэробные микроорганизмы из рода Pscudomonas и Paracoccus, а также термофильные спорообразующие микроорганизмы. Восстанавливать нитраты могут некоторые хемолитоавтотрофы, типичным представителем которых является Thiobacilius denitrificans. Этот микроорганизм размножается в анаэробных условиях, используя нитраты в качестве акцептора водорода.
Диссимиляционная денитрификация наносит большой ущерб сельскому хозяйству, так как приводит к снижению запасов азота в почве.
Косвенная денитрификация - это чисто химический процесс взаимодействия нитритов с аминокислотами, в результате которого образуется молекулярный азот:
R-CHNH2-COOH + HNO2 R-CHOH-COOH + N2 + H2O
Дата публикования: 2015-11-01; Прочитано: 2999 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
