 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Строение бактериальной клетки
|
|
Основной физической единицей всего живого является клетка. Энергетические реакции, природа генетического материала, аппарат его репликации и механизм биосинтеза белка во всех клетках, в основном, одинаковы. Но прокариотическая бактериальная клетка обладает рядом характерных особенностей, которые отличают ее от эукариотической клетки. Среди этих особенностей следует отметить тип рибосом (у бактерий рибосомы 70S - типа), морфологическую организацию ядра и аппарата дыхания (у бактерий нет митохондрий), отсутствие эндоплазматической сети, неспособность бактерий синтезировать стерины и наличие особых гетеро-полимеров, входящих в состав клеточной стенки.
У эукариотов имеется истинное ядро, отделенное от цитоплазмы ядерной мембраной. В ядре содержится набор хромосом, который после удвоения в процессе митоза передается двум дочерним клеткам. Цитоплазма эукариотической клетки содержит митохондрии и пластиды. Цитоплазм этическая мембрана переходит внутри клетки в эндоплазм этическую сеть и в ядерную мембрану-
Прокариоты представляют собой, в основном, одноклеточные, но также нитчатые, мицелиальные и колониальные организмы, лишенные хлорофила и обитающие во влажных средах.
Прокариоты являются реликтовыми формами живых организмов, сохранившихся с древнейших, времен. Основным отличительным признаком прокариотов является отсутствие ядра окруженного мембраной. Вместо ядра у бактерий имеется образование, получившее название «нуклеоид». Нуклеоид бактерий состоит из нуклеопротеидов, основным компонентом которых является ДНК. Он не ограничен от цитоплазмы какой-либо ядерной мембраной и не связан с основным протеином бактериальной клетки.
Клеточное деление прокариот не сопровождается циклическими изменениями в консистенции или в способности к окрашиванию пи нуклеоплазмы, ни цитоплазмы. Во время клеточного деления в цитоплазме прокариот не образуется система микротрубочек веретена. У них наблюдаются процессы переноса и рекомбинации генов, но никогда не происходит гаметогенез и формирование зигот.
Цитоплазматическая мембрана прокариот отличается сложным строением и образует различные инвагинации в цитоплазму. Другой особенностью прокариот является отсутствие цитоплазматических органелл, например, митохондрий, окруженных элементарной мембраной. Вакуоли и репродуцирующие цитоплазматические органеллы встречаются сравнительно редко и окружены мембранами.
Цитоплазма прокариот неподвижна: тока цитоплазмы, псев-доподиальных движений, эндоцитоза и экзоцитоза нет. Питательные вещества поглощаются в молекулярной форме.
У большинства прокариот клетка окружена ригидной клеточной стенкой, не универсальной по составу и строению. Она состоит из гетеро поли мерно го вещества, которое не было обнаружено ни у одного из эукариотических организмов.
Клетки прокариот могут быть неподвижными или обладать способностью к «плаванию», либо к скользящему движению по поверхности. Органы движения прокариот значительно упрощены и имеют принципиально иное строение, чем у эукариотов.
К числу основных компонентов бактериальной клетки относятся: клеточная стенка, цитоплазм этическая мембрана, цитоплазма и нуклеоид (рис. 7).
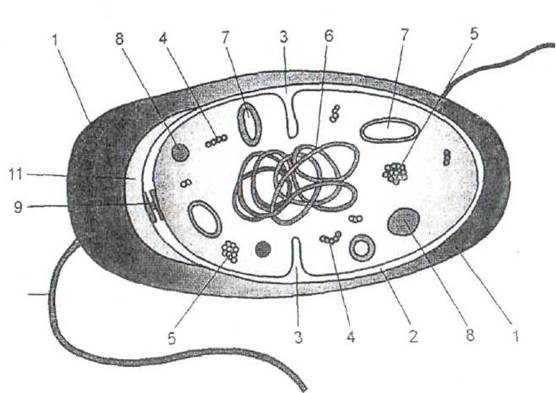
Рис. 7. Строение микробной клетки
1- клеточная стенка; 2 - цитоплазматическая мембрана; 3 - мезосомы; 4 – рибосомы;
5 - полисомы; 6 - клубок кольцевой хромосомы, состоящей из молекулы ДНК (нуклеоид);
7 - плазмида; 8 - запасные вещества и внутриклеточные включения;
9 - диски базального тела жгутика; 10 - жгутик; 11 - периплазма
Клеточная стенка. Под клеточной стенкой понимают образование, расположенное снаружи бактериальной клетки и окружающее цитоплазматическую мембрану. Основная функция клеточной стенки состоит в защите цитоплазмы и цитоплазматической мембраны от механического повреждения и неблагоприятного влияния факторов внешней среды. Кроме того, она играет существенную роль в процессе роста и делении клеток.
Клеточная стенка проницаема для солей и других многочисленных низкомолекулярных соединений. Она регулирует приток воды внутрь клетки и внутреннее гидростатическое давление. Высокая внутриклеточная концентрация метаболитов обусловливает значительный перепад осмотического давления между цитоплазмой и средой обитания бактерий. Внутреннее осмотическое давление у большинства микроорганизмов колеблется от 5атм. до 20 атм. Прочная и эластичная клеточная стенка предотвращает разрыв клетки и позволяет ей сохранять определенную форму.
Химический состав и строение клеточной стенки постоянны для определенного вида микроорганизмов. Основным материалом клеточной стенки является сложный полимер пептидогликан. Важной особенностью пептидогликана является его прочность, эластичность, способность задерживать нежелательные для клетки химические вещества, в том числе антибиотики.
Пептидогликановые цепочки связаны между собой многочисленными перекрестными связями, образуя одну гигантскую молекулу. Этим достигается механическая прочность бактериальной клетки.
Длина цепочек пептидогликана, а также способ и частота их соединения определяют форму бактериальных клеток.
В зависимости от строения клеточкой стенки прокариоты делят на две группы: грамположительные и грамотрицательные. Установлено, что грамположительные бактерии после окрашивания основными красителями три фенил метанового ряда (кристалвиолетом или генцианвиолетом) и обработки иодом, утрачивают способность обесцвечиваться спиртом. Аналогичная окраска грамотрицательных бактерий не приводит к образованию прочного окрашенного комплекса. Они легко обесцвечиваются спиртом или ацетоном,
Клеточные стенки грамположительных и грамотрицательных бактерий различаются по своему химическому составу и ультраструктуре. У грамположительных микроорганизмов имеется несколько пептидогликановых слоев, на которые приходится от 50% до 90% веса клеточной стенки. У грамотрицательных бактерий содержание пептидогликана в клеточной стенке колеблется от 1% до 10 %.
С помощью электронного микроскопа показано, что клеточная стенка грамположительных бактерий устроена менее сложно и выглядит как гомогенный толстый пептидогликановый слой. У грамотрицательных бактерий клеточная стенка состоит из липополисахарида, наружной мембраны и л и поп роте и на, которые трехконгурным слоем расположены над пептидогликаном. Ли по протеин связывает пептидогликан с наружной мембраной, а липололисахарид крепится к наружной мембране с помощью гидрофобных связей (рис. 8). Такой комплекс создает дополнительный барьер на пути проникновения в клетку различных веществ. Поэтому грамотрицательные бактерии более устойчивы к воздействию различных химических, в том числе и дезинфицирующих, веществ.
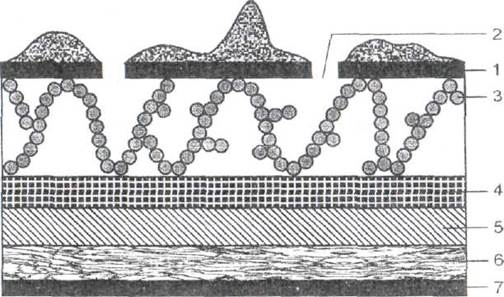
Рис. 8. Схематическое строение клеточной стенки грамотрицательных бактерий
1 - липополисахаридный слой с липопротеидными бугорками;
2- каналы липополисахарвдного слоя; 3 - молекулы белка (гидрофобные связи);
4 - наружная мембрана; 5 - липопротеин; 6 - пептидогликан; 7 - цитоплазматическая мембрана.
Некоторые микроорганизмы, подвергаясь воздействию различных неблагоприятных факторов, могут лишаться клеточной стенки. Это происходит под влиянием лизоцима, который разрушает клеточную стенку, в результате мутации или блокирования синтеза пептидогликана антибиотиками и токсичными агентами.
Бактерии, полностью лишенные клеточной стенки, приобретают шаровидную форму и называются протопластами. Появление протопластов характерно для грамположительных бактерий. У грамотрицательных бактерий клеточная стенка, как правило, полностью не удаляется. Поэтому при разрушении клеточной стенки грамотрицательные бактерии превращаются в сферопласты.
Протопласты и сферопласты являются весьма неустойчивыми структурами. Они очень чувствительны к изменениям осомотического давления, к механическим воздействиям, не подвергаются инфицированию бактериофагами и не обладают активной подвижностью.
Если протопласты или сферопласты сохраняют способность к росту и размножению, то их называют L-формами бактерий. Свое название они получили в честь института им.Листера в Лондоне, в котором они были открыты в 1935 году Различают нестабильные и стабильные L-формы. Первые сохраняют свои свойства только в присутствии фактора, который вызвал разрушение клеточной стенки. После устранения этого фактора наблюдается реверсия, т.е. восстановление клеточной стенки и формы микроорганизма. Стабильные L-формы уже никогда не подвергаются реверсии.
У некоторых бактерий снаружи клеточной стенки откладывается более или менее толстый слой сильно обводненного слизистого материала. Если этот материал плотно облегает клеточную стенку и практически неразделим с ней, то образуется чехол. При свободном облегании материала образуется капсула, а если материал слабо связан с клеточной стенкой, то это определяется как слизистый слой.
В зависимости от толщины и строения различают микро- и макрокапсулы. Капсульное вещество бактерий состоит из полисахаридов, мукополисахаридов или полипептидов. Оно имеет аморфное строение и в различной степени сохраняет связь с клеточной стенкой. Роль капсулы точно не определена, но считают, что капсулы не имеют жизненно важного значения для микроорганизмов. Но капсулы предохраняют клетку от высыхания, повышают устойчивость бактерий к фагоцитозу, предохраняют от иммунных механизмов инфицированного организма и неблагоприятных факторов внешней среды. Капсулы могут играть роль ионообменника, поглощая питательные вещества и освобождая их впоследствии для использования клеткой.
Капсулы могут образовываться не при всех условиях окружающей среды, могут появляться мутанты, не обладающие способностью к их образованию. С помощью ферментов слизистый слой можно удалить, не причиняя вреда бактериям.
У некоторых сапрофитных организмов (лейконосток) наблюдается образование общей капсулы для многих клеток. Такое скопление микроорганизмов, заключенных в общую капсулу, называют зооглеями (рис. 9).
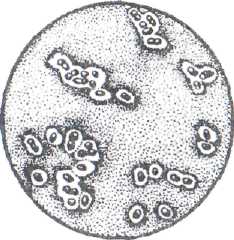
Рис.9. Скопление микроорганизмов, заключенных в общую капсулу (зооглеи)
Между клепочной стенкой и цитоплазматической мембраной располагается перизматическое пространство (периплазма), содержащее ферменты.
Цитоплазматическая мембрана располагается между клеточной стенкой и цитоплазмой. Небольшая толщина цитоплазматической мембраны не позволяет ей выступать в роли пластического материала, но она играет ведущую роль в процессах переноса молекул из питательного раствора в клетку. Цитоплазматическая мембрана представляет собой сложное высокоорганизованное и высокоспециализированное образование, состоящее из трех слоев: липидного, протеинового и полисахаридного. В ее состав входят белки, фосфолипиды, липопротеин, небольшое количество углеводов и другие соединения. Цитоплазматическая мембрана выполняет функцию разделительной перегородки между клеточной стенкой и содержимым клетки. Она обеспечивает постоянство внутриклеточного осмотического давления и высокую избирательную проницаемость для различных веществ, регулируя их поступление в бактериальную клетку и выведение из клетки продуктов метаболизма. Активное продвижение через цитоплазм этическую мембрану различных веществ и ионов, необходимых для жизнедеятельности клетки, непрерывно осуществляется только с помощью ферментов (пермеаз). Поэтому цитоплазматическая мембрана играет важную роль в обмене веществ. Ионизированные молекулы, такие, как органические кислоты, аминокислоты, неорганические соли, плохо проникают через цитоплазматическую мембрану, т.к. они отталкиваются электрическим зарядом поверхности мембран, а также потому, что мембрана не имеет достаточно больших пор для прохождения крупных молекул. Это определяет барьерные функции цитоплазматической мембраны и ее избирательность по отношению к веществам, окружающим клетку.
В цитоплазматической мембране локализованы высокочувствительные рецепторы, с помощью которых клетки распознают и обрабатывают сигналы, поступающие из окружающей среды, дифференцируют питательные вещества и различные антибактериальные соединения. На поверхности цитоплазматической мембраны содержатся активные ферментные системы, принимающие участие в синтезе белка, токсинов, ферментов, нуклеиновых кислот и других веществ. Прокариотная мембрана играет также важную роль в дыхании клетки, в процессах окислительного фосфорилирования.
Цитоплазматическая мембрана участвует в процессах синтеза компонентов клеточной стенки и капсулы, в делении нуклеоида, в формировании эндоспор.
При вдавливании (инвангинации) в сторону цитоплазмы цитоплазматическая мембрана образует мезосомы (от греч. mesos - средний и soma - тело), на которых фиксируется ДНК. Мезомомы содержат большое количество ферментов и играют важную роль в формировании клеточных стенок, делении, дыхании. Некоторые ученые считают мезосомы аналогами митохондрий, так как они связаны с ферментами цитохромной системы окислительного фосфорилирования.
Цитоплазма бактерий представляет собой коллоидную систему, отделенную мембраной и состоящую из воды, белков, углеводов, липидов, минеральных соединений и других веществ. В отличие от животных и растительных клеток, цитоплазма бактерий не содержит органелл. Кроме того, у бактерий цитоплазма ограничена только снаружи, тогда как в клетках высших организмов она отграничена изнутри ядерной мембраной. 11ри старении клетки дисперсность цитоплазмы изменяется, она принимает ячеистое строение с образованием вакуолей, наполненных клеточным соком.
Бактериальная цитоплазма пронизана многочисленными структурами, неподвижна и имеет высокую плотность. Она содержит мелкие включения, состоящие на 60% из РНК и на 40% из протеина. Эти включения получили название рибосом. При активном синтезе белка из нескольких рибосом образуются агрегаты, называемые полисомами или полирибосомами. Полисомы осуществляют синтез белка и являются весьма важными структурами для жизнедеятельности бактерий.
В цитоплазме находятся различные включения, которые представлены полисахаридами (крахмал, гликоген), липидами, полипептидами, полифосфатами, пигментными скоплениями, отложениями серы и кальция. Они откладываются в цитоплазме в условиях избытка питательных веществ в окружающей среде.
Некоторые микроорганизмы обладают способностью запасать фосфорную кислоту в виде гранул полифосфата или волютина. Эти гранулы используются клетками как источник фосфора. Наличие гранул волютина является специфическим признаком, который учитывают при лабораторной диагностике, например, дифтерийных бактерий.
Жировые скопления, или липопротеиновые тельца, довольно часто встречаются у бацилл и спирилл. Они появляются при росте клеток на питательных средах, содержащих много углеводов, и исчезают при голодании. Биологическое значение зерен волютина и липопротеиновых включений состоит в том, что они служат запасным питательным веществом и используются бактериями при голодании.
В цитоплазме бактерий имеются вакуоли, которые состоят из различных, растворенных в воде веществ, и окружены мембраной (тонопласт) липопротеидного происхождения. Число вакуолей в клетке колеблется в пределах 6-10, а в период активного роста увеличивается до 20.
Нуклеоид бактерий представляет собой морфологическую структуру, которая отличается от ядер растительной и животной клетки отсутствием оболочки, ядрышка, а также цитотического способа деления. Генетический материал нуклеоида сосредоточен в одной бактериальной хромосоме, имеющей форму замкнутого кольца и расположенной в центральной зоне клетки. ДНК нуклеоида содержит наследственную информацию и руководит всей деятельностью клетки.
Кроме хромосомы нуклеоида, в цитоплазме расположены ко-валенгные замкнутые кольца ДНК, так называемые плазм иды. Они придают бактериям способность к коньюгации, устойчивость к антибиотикам, токсигенные свойства и др.
Органеллы движения. Движение бактерий - одно из самых поразительных явлений природы, которое впервые наблюдал А. Левенгук. Современные исследования показали, что бактерии, используют для своего движения устройство, куда более сложное, чем колесо, а именно ротор, вращающийся под действием электрического поля. По существу речь идет об электродвигателе субмикронного размера, собранном из нескольких белковых молекул.
Благодаря такому электродвигателю многие виды бактерий способны плыть сквозь толщу воды со скоростью до 150 мкм в секунду, то есть за одну секунду бактерия покрывает расстояние, примерно в сто раз превышающее ее длину.
Основным двигательным органоидом бактерий служат жгутики (флагеллы). Жгутики представляют собой специальные нитевидные образования, состоящие из одного единственного белка - флагеллина -молекулярной массой от 20 килодальтон до 60 килодальтон (kDa), в зависимости от вида бактерии (рис. 10). Белок жгутиков обладает антигенными свойствами. Бактериальный жгутик имеет в поперечнике всего около 0,01 мкм, а его длина может в 10 раз превышать длину самой бактерии.
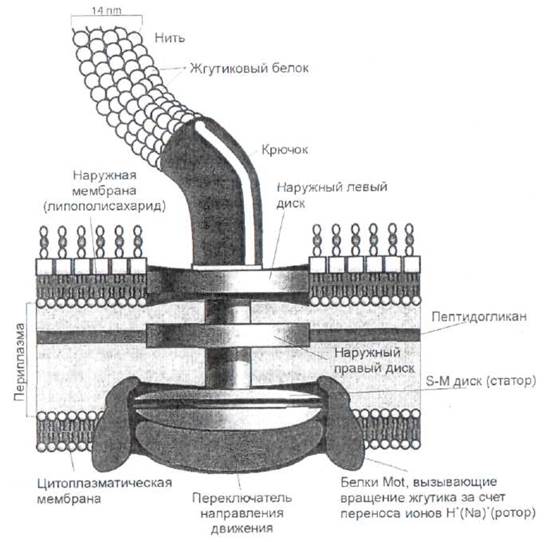
Рис. 10. Строение жгутика грамотрицательных бактерий.
Работы последних лет показали, что бактериальный жгутик вращается вокруг длинной оси. Источником энергии для вращения служит электрическое поле или ионный градиент, создаваемый потоком заряженных частиц: ионов Н+ или Na+, перемещающихся через клеточную мембрану из окружающей среды внутрь клетки.
Жгутик бактерии представляет собою жесткую полую трубку в форме, как правило, левой спирали длиной несколько микрон (иногда до 20), крепящейся к дискам специального образования - базального тела.
Наружный диск находится в клеточной стенке, а внутренний - в цитоплазматической мембране.
Вращение дисков относительно друг друга сообщается жгутикам. При движении бактерии ее жгутик совершает вращение вокруг своей оси по часовой стрелке, либо - против. В результате энергичного движения жгутиков, напоминающего вращение штопора, жидкость движется вдоль жгутиков, и бактерии передвигаются.
У некоторых бактерий есть всего один жгутик, у других - их может быть до нескольких десятков. Расположение жгутиков у подвижных бактерий - характерный признак, который имеет таксономическое значение. В зависимости от расположения жгутиков, микробы подразделяются на четыре группы (рис. 11):
1. Монотрихи - бактерии с одним жгутиком на конце (холерный вибрион, синегнойная палочка).
2. Лофотрихи - бактерии, которые имеют по пучку жгутиков на одном конце.
3. Амфитрихи - бактерии с двумя полярно расположенными жгутиками или имеющие по пучку жгугиков на обоих концах.
4. Перитрихи - бактерии, обладающие жгутиками по всей поверхности тела (кишечная палочка, сальмонеллы).
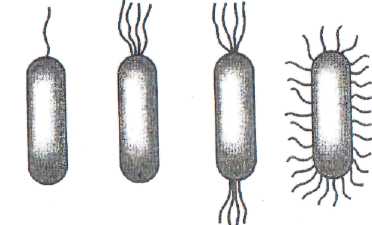
Рис. 11. Разновидности жгутиковых бактерий
1 - монотрихи; 2 - лофотрихи; 3 - амфитрихи; 4 - перитрихи.
Подвижные бактерии способны к направленным движениям, или таксисам. В зависимости от факторов среды, под влиянием которых происходит движение, различают хемотаксис, аэротаксис и фототаксис. При хемотаксисе бактерии уходят из зоны высокой или низкой концентрации веществ в места более благоприятного обитания. Аэротаксис определяется отношением бактерий к кислороду, а фототаксис - отношением бактерий к солнечному свету как источнику энергии.
Фимбрии. У различных видов микробов имеются фимбрии (реснички, пили, филаменты), представляющие собой образования значительно короче и тоньше жгутиков, Они покрывают тело клетки. Количество их достигает от 100 до 400 на одной особи. Полагают, что они не служат для передвижения, а способствуют прикреплению клеток к поверхности различных субстратов. Некоторые пили (F-пили) осуществляют направленный половой процесс передачи генетического материала из клетки-донора (мужской штамм) в клетку-реципиент (женский штамм). Такой процесс называется конъюгацией.
Способность клетки быть донором связана с наличием особого генетического фактора, который при коньюгации переносится из одной клетки в другую. Этот генетический фактор носит название мужского полового фактора или фактора F (от англ. fertility - плодовитость). F фактор переносится из одной клетки в другую с помощью половых волосков или F-пилей при непосредственном контакте клеток. При этом женские клетки всегда становятся мужскими.
F фактор не содержится в хромосомах, он относится к вне-хромосомным ДНК - плазмидам. Многие плазмиды несут гены, ответственные за процесс коньюгации. К ним относятся гены, определяющие некоторые поверхностные структуры клетки. При соприкосновении бакгериальных клеток эти структуры участвуют в образовании коньюгационного мостика.
Таким образом, коньюгация представляет собой процесс, способность к которому обеспечивается плазмидой и результатом которого является перенос плазмидной ДНК. Гены бактериальной хромосомы переносятся в клетку-реципиент, если они интегрированы с плазмидой в донорской клетке.
С помощью коньюгации из клетки в клетку передаются факторы передачи устойчивости Rtf (от англ. resistance transfer factors - факторы передачи устойчивости). Они содержат гены, которые передают бактерии-хозяину устойчивость к антибиотикам, химиопрепаратам, температуре и другим факторам окружающей среды. Например, приобретенная устойчивость микроба к воздействию химического вещества передается из мужской клетки в женскую с помощью F-пиля.
Дата публикования: 2015-11-01; Прочитано: 7178 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
