 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Раковины фораминифер
|
|
|
|
 а
а
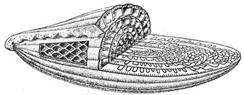
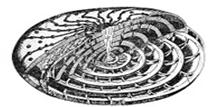
Рис. 23. Известковые раковины фораминифер:
Нуммулитиды (K 2 – Q): а – внутреннее строение раковины нуммулитид,
б – скопление раковин современных нуммулитид.
Фузулиниды (C – P): в – внутреннее строение раковины фузулинид.
фораминифер являются бентосными организмами, но на рубеже пермского и триасового периодов часть из них приспосабливается к планктонному образу жизни. В позднемеловую эпоху планктонные фораминиферы получают особенно широкое распространение (отряды Глобигериниды и Глобороталииды). Появление планктонных фораминифер, по мнению американских исследователей, могло стать реакцией на потепление и понижение содержания кислорода в океанической воде.
Класс Радиолярии(Radiolaria) – морские планктонные организмы, размеры которых варьируют от нескольких десятков микрон до 1-2 мм, а колониальные формы достигают нескольких дециметров. В целом, однако, радиолярии мельче фораминифер. Важным элементом строения радиолярий является минеральный скелет, состоящий из аморфного кремнезема или целестина.
А б
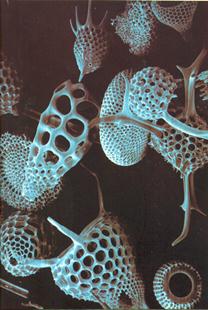
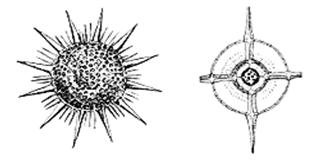
Рис. 24. Радиолярии:
(а, в) – скелеты радиолярий
б – поперечное сечение скелета радиолярий
|
Кремнистый скелет сохраняется в ископаемом состоянии, целестиновый – растворяется после смерти организма. Скелет имеет сферическую, звездчатую, шлемообразную или колоколообразную форму (рис. 24 а,в). Он состоит из вложенных один в другой решетчатых элементов и длинных радиальных игл (спикул), отходящих от центра (рис. 24 б). По особенностям строения скелета разные авторы выделяют от 2 до 5 отрядов радиолярий. Скелет частично выполняет защитные функции, но главное – увеличивает поверхность тела, обеспечивая парение в толще воды.
Появление радиолярий датируется ранним палеозоем, но есть данные о находках этих организмов в позднем докембрии. В современных морях радиолярии распространены в низких широтах, где накапливаются их скелетные остатки, образуя радиоляриевые илы. Стратиграфическую значимость радиолярии получают, начиная с мезозоя. В настоящее время насчитывается не менее 7000 видов этого класса, в то время как в ископаемом состоянии известно около 1000.
2.2.3.2. Подцарство Многоклеточные(Metazoa)
В отличие от простейших тело многоклеточных состоит из значительного количества клеток и межклеточного вещества, образующих ткани и органы. Выделяется 4 типа тканей, различающихся строением и функциями: эпителиальная, соединительная, мышечная и нервная. Органы многоклеточных, состоящие из комплекса тканей, регулируются нервной и кровеносной системами. Для многоклеточных характерно половое размножение, но у примитивных (низших) многоклеточных может наблюдаться чередование полового и бесполого.
Надраздел Низшие многоклеточные (Parazoa)
К примитивным многоклеточным относятся губковые, археоциаты и некоторые группы организмов неясного систематического положения (Приложение 3).
В состав типа Губковые (Spongia) входят морские и пресноводные прикрепленные бентосные организмы, тело которых состоит из 2 слоёв. Между слоями находится желеобразное вещество мезоглея, в которой формируется скелет, состоящий из иголочек (спикул). По способу питания губковые фильтраторы. Тело пронизано системой каналов, по которым поступает вода, обеспечивающая поступление пищи. Устья каналов (поры) обусловили другое название типа – Porifera (Пороносцы). До середины ХХ века в состав порифер включались археоциаты, позднее выделенные в особый тип. В последние годы появляются данные о целесообразности объединения губок и археоциат в тип Porifera. Согласно современной классификации Губковые подразделяются на три класса, из которых наибольшее распространение (около 10000 видов) имеет класс Губки. Остальные два класса (Коралловые губки и Сфинктозоа) встречаются сравнительно редко и в данной работе не рассматриваются.
По химическому составу скелета класс Губки расчленяется на подклассы Кремневые (Опаловые) и Известковые губки. Более дробная классификация основывается на структуре спикул. В ископаемом состоянии кремневые губки известны с кембрия, а известковые – с девона. Имеются указания о находках спикул в отложениях венда. Известковые губки участвуют в формировании рифов, кремневые губки современных осадков в процессе диагенеза могут превращаться в яшмы,  трепелы, опоки.
трепелы, опоки.


А б в
Рис. 25. Тип Губковые: а – вымершая одиночная губка (K),
б – колония вымерших губок,
в – современная морская губка
Тип Археоциаты (Archaeocyathi) (С) – вымершие бентосные (обычно прикрепленный бентос) одиночные и колониальные организмы губкоподобного облика. Наличие пор, в которые поступала вода, и центрального отверстия, через которое она выливалась, сближает археоциат с губками. Однако, в отличие от губок, скелет археоциат был не спикульный, а зернистый и только известковый. Одиночные археоциаты имели кубкообразную, реже цилиндрическую или блюдцеобразную форму. Диаметр кубка составлял 1-5 см, высота – 6-10 см. Колониальные археоциаты имели ветвистый, иногда массивный облик. Скелет археоциат обычно состоял из внутренней и наружной стенок, в промежутке между которыми располагалось тело организма, заполненное вертикальными перегородками (класс Правильные) илистерженьками (класс Неправильные) (рис. 26 1,2).
При благоприятных условиях скопления археоциат превращались в древнейшие рифы (рис. 27). Археоциаты являлись одной из быстро эволюционирующих групп морской фауны. Они появились в начале раннекембрийской эпохи, достигли расцвета и вымерли до её окончания. Не исключено, что вымирание археоциат на ранне-среднекембрийской границе было, в некоторой мере, кажущимся.
Возможно известковые губки, достоверно известные с девонского периода, были генетически как-то связаны с археоциатами. В настоящее время найдена губка Vacletia, морфологически сходная с археоциатами. На современном уровне знаний очевидна стратиграфическая значимость археоциат для раннекембрийской эпохи.
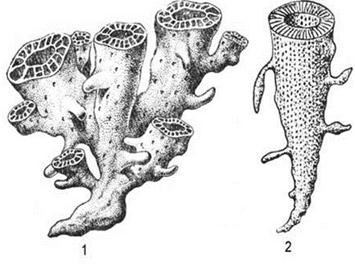
Рис. 26. ТипАрхеоциаты:
1 – колония археоциат, 2 – одиночная форма археоциат

Рис. 27. Рифовый известняк, образованный скелетами археоциат
Надраздел Настоящие многоклеточные (Высшие) (Eumetazoa)
Настоящие многоклеточные объединяют организмы, характеризующиеся дифференцированными тканями и органами. Они развиваются из двух или трех зародышевых листков (экто-, эндо- и мезодермы), соответственно, подразделяясь на радиально- и двусторонне-симметричных (Приложение 4).
Раздел Радиально-симметричные (Radiata)
Радиально-симметричные организмы (радиалии) обычно принадлежат к прикрепленному бентосу или планктону. Из ископаемых представителей радиалий наибольшее распространение имеет тип Стрекающие.
Стрекающие (Cnidaria) обитают исключительно в водной среде, хотя некоторые виды могут находиться на суше во время отлива. Лишь небольшая часть животных приспособилась к жизни в пресных водоемах (гидры и гидромедузы). Уникальная черта стрекающих — наличие стрекательных клеток, которые они используют для охоты и защиты от хищников, что и послужило поводом для названия этого типа. Каждая стрекательная клетка снабжена чувствительным волоском, который отвечает за выброс стрекательной нити, через которую в организм жертвы и попадает яд (рис. 28). Описано около 11 тысяч видов современных и вымерших книдарий.
Жизненный цикл стрекающих имеет две стадии: свободно плавающую форму медузы и прикрепленную полипов (рис. 29). Многие виды чередуют бесполую стадию полипа и половую медузы, у некоторых цикл лишён одной из стадий. Тело книдарий обладает радиальной симметрией и представляет собой открытый на одном конце двухслойный мешок (кишечную полость), другой конец замкнут. Единственное отверстие, ведущее в полость тела (рот), используется для дыхания, пищеварения и выделения. Вокруг рта располагается венчик щупалец со стрекательными клетками. Все книдарии хищники, имеют примитивную нервную систему, состоящую из отдельных нервных клеток.
Многие виды книдарий образуют колонии (обычно путём почкования), являющие собой единый организм, состоящий либо из медузоподобных, либо полипоподобных особей. В колонии все особи находятся в органической связи друг с другом, осуществляя общий обмен веществ, направленный на поддержание жизнедеятельности колонии. У колониальных полипов, кроме того, развивается общий скелет. Во время полового размножения в колонии образуются мужские и женские половые клетки, которые выпускаются в воду, где и происходит оплодотворение. Образуется личинка – планула, которая является расселительной формой. Она некоторое время плавает, затем прикрепляется к субстрату, давая начало полипу (рис. 29).
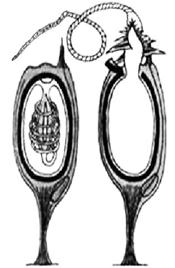
Рис. 28. Стрекательные клетки книдарий:
а – стрекательная клетка в покое,
б – стрекательная клетка в действии
А б
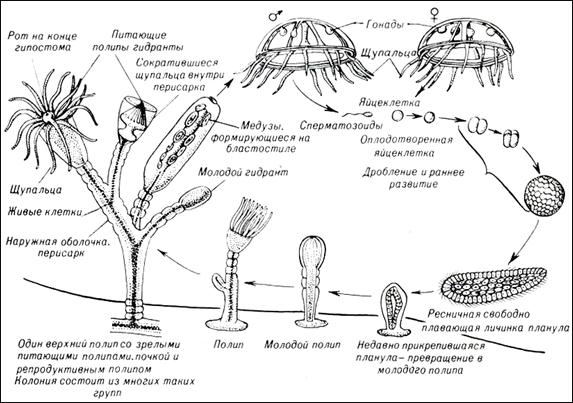 Рис. 29. Схема полового размножения полипов.
Рис. 29. Схема полового размножения полипов.
В настоящее время тип насчитывает 7 классов, из которых для палеонтологии важны два: Гидроидные (Hydrozoa) и Коралловые полипы (Anthozoa).
К классу Гидроидные (Hydrozoa) относятся наиболее примитивные стрекающие, чаще колониальные, реже одиночные с радиальной или четырехлучевой симметрией тела. Вокруг рта располагается 1-3 венчика щупалец, которые используются, в основном, для захвата пищи. Брюшная полость не разделена на перегородки. Скелет хитиноидный или карбонатный. Размножение бесполым путем (почкование) или половым (чередование полипоидной и медузоидной стадий). В отличие от других классов эти стадии носят названия гидрополипов и гидромедуз. В ископаемом состоянии гидроидные известны с кембрия. У некоторых представителей этого класса, например, у гидры пресноводной, развита только полипоидная стадия. Морские гидроидные основной жизненной формой имеют медузу. Для стратиграфии важна группа пока до конца неясного систематического происхождения — строматопораты.
Подкласс Строматопораты (Stromatoporata) традиционно относят к Hydrozoa. Однако в современных классификациях их часто сближают с губками и археоциатами. Это вымершие колониальные организмы; от них сохранились известковые слоистые скелеты разнообразной формы: от пластинчатой, корковидной до ветвистой (рис. 30). Слоистые скелеты представляют собой наслаивающиеся параллельно друг на друга пластины — ламины. В шлифах при увеличении видно, что ламины имеют разнообразное строение: пузырчатое, губчатое, петельчатое и т.д. Скелет строматопорат пронизан разнообразными пустотами. Самыми примечательными из них являются астроризы — каналы, не имеющие самостоятельных стенок, зато имеющие на верхней поверхности ламин вид звездчатых пустот, обычно с длинными горизонтальными разветвлениями (рис. 31). Они сходны со звездчатыми каналами некоторых губок, но также напоминают циклосистемы гидрокораллов. Отсюда дискуссионность их систематического положения: тип губки (Spongiata) или тип стрекающие (Cnidaria). Перпендикулярно ламинам располагаются зубчики, пересекающие ламины, шипики и стерженьки — столбики (рис. 32). Зубчики, шипики и стерженьки располагаются изолированно или создают различные комплексы в виде щеток, сеток, петель, трубок, колонн и пр. Строматопораты — донные колониальные животные, прикрепленные или свободнолежащие, нередко входившие в состав банок и рифов, образующие в ископаемом состоянии строматопоровые известняки. В палеозое (поздний ордовик, силур и девон) и мезозое (триас и юра) строматопораты активно участвовали в рифостроении. В отложениях каменноугольного и пермского периодов строматопораты практически не встречаются. Они существовали в нормально-морских условиях, но могли выдерживать и колебания солености, так как их находки известны вдоломитах. Форма палеозойских колоний разнообразная: пластинчатая, корковидная, полусферическая,


Рис. 30. Ископаемые колонии строматопорат


Рис. 31. Астроризы (звездчатые каналы) Рис. 32. Ламины с вертикальными
шипиками и зубчиками
дендроидная. Для мезо-кайнозойских строматопорат характерны сферические колонии.
Класс Коралловые полипы (Anthozoa) по форме тела и яркой окраске напоминают цветы, anthos — цветок. Это исключительно морские книдарии, требовательные к нормально морским условиям среды. Малейшее изменение температуры солености и газового режима грозит им гибелью. Коралловые полипы могут быть колониальные (рис. 33,а), или одиночные (рис. 33,б), и те и другие имеют только полипоидную стадию существования. В отличие от гидроидных коралловые полипы имеют более высоко организованное строение гастральной полости. Она разделена перегородками (мезентериями) на центральную пищеварительную полость и периферические камеры. Вокруг рта расположены щупальца (от 8 до нескольких десятков). Коралловые полипы (Anthozoa) могут питаться двумя способами. Первый основан на симбиозе с одноклеточными водорослями – зооксантеллами, обитающими в тканях животных. Водоросли получают от хозяина азотистые соединения, фосфаты и углекислоту, отдавая взамен полученные в результате фотосинтеза аминокислоты, углеводы и главный энергетический материал кораллов – липидные соединения. Второй (хищный) способ предусматривает захват из окружающей среды различных организмов.
Скелет кораллов – кораллит может быть роговой или карбонатный, (наружный или внутренний), некоторые виды вообще не имеют скелета. Кораллы обитатели морей всех глубин от литорали до ультраабиссали. Первые коралловые полипы известны из кембрия.


Рис. 33.а Колония коралловых полипов Рис. 33.б Одиночные полипы
В ископаемом состоянии скелеты кораллов образовывали органогенные постройки – биостромы и биогермы. Биостром – плоская органогенная постройка, из нарастающих друг на друга рифообразователей, не достигающая уровня воды. В ископаемом состоянии она выглядит как линза массивного известняка среди слоистых известняков. Разновидностью биострома является банка. Биогерм – изометричная органогенная постройка, резко возвышающаяся над морским дном, достигающая поверхности воды. В ископаемом состоянии это массивный известняк, «протыкающий» слоистые образования вокруг него. Риф (рис. 34) является разновидностью биогерма. По форме и расположению относительно берега рифы делятся на примыкающие (являются продолжением берега), барьерные (отделяют прибрежную лагуну от открытого моря) и атоллы (подводные горы, увенчанные рифовой постройкой) (рис. 34 а, б, с). Вокруг рифа (зона риффлет, рис. 34, В), накапливаются продукты его разрушения – глыбы, дресва, щебень, которые в процессе диагенеза превращаются в известковую брекчию. Высота рифов колеблется от нескольких метров до сотен метров. Древние коралловые рифы являются «ловушками» для нефти. Многие разрабатываемые нефтяные месторождения связаны с рифами.
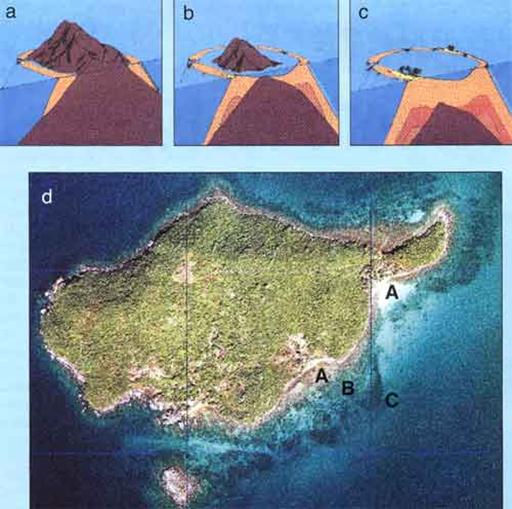
Рис. 34. Типы и зональность рифов:
a - примыкающий, b - барьерный, c - атолл,
Различные зоны: A - лагуна, B - риффлет, C - склон рифа
В состав банок и рифов, как правило, входят скелеты не только кораллов, но и других организмов: моллюсков, брахиопод, губок. В строительстве рифов у кораллов есть помощники — определенные виды морских водорослей, скрепляющих коралловую массу.
Современный коралловый риф – это сложная и хрупкая экосистема, изобилие разных, притом чрезвычайно тесно соседствующих существ. Дополняют и усиливают эту экстраординарную интенсивность жизни сказочная красота форм и разнообразие красок. На коралловом рифе в одном месте и в одно время можно увидеть представителей практически всех структурных уровней развития организмов. Здесь представлен полный их спектр: от сине-зеленых морских водорослей и бактерий, многочисленных типов беспозвоночных животных до рыб, рептилий, птиц и млекопитающих.
Современные рифы заметно страдают от прожорливых врагов — морских звезд. Морская звезда «терновый венец» способна за день уничтожить тысячи полипов, оставляя хрупкие белые скелеты. Эти хищники нанесли немалый урон коралловым рифам у Японских берегов, а также Большому Барьерному рифу Австралии. В настоящее время выяснилось, что у морских звезд есть враг — гигантский моллюск тритон, который активно их уничтожает. Оказалось весьма эффективным в качестве охранных мероприятий выращивать тритонов в специальных питомниках и выпускать их на рифы.
Подкласс Табулятоморфы (Tabulatomorpha) объединяетвымерших палеозойских исключительно колониальных коралловых полипов, которые жили с позднего кембрия и вымерли в карбоне. Скелет их состоял из небольших (0,5-4 мм) трубчатых кораллитов, округлых или многоугольных в сечении (рис. 35; а, б).

А б в
Рис. 35. Строение кораллитов табулятоморф:
а – общий вид, б – поперечное сечение,
в – продольное сечение с днищами, п – поры
Внутренние элементы кораллита – горизонтальные или воронковидные днища и радиальные септальные шипики (рис. 35, в; 37, 3а). Кораллиты были соединены друг с другом порами или соединительными трубочками; у некоторых видов связь между кораллитами отсутствовала. Колониимассивные, цепочечные (рис. 37, 1,2,3) кустистые, или стелющиеся.
Скелет по составу карбонатный. Подкласс подразделяется на отряды: аулопориды, хализитиды, фавозитиды, михелениды, сирингопориды и другие. От аулопорид возникли ругозы. Табулятоморфы обитали в мелководной зоне морей и входили в состав рифов. Имеют значение для стратиграфии палеозойских отложений.
Подкласс Ругозы ( Rugosa) или четырехлучевые кораллы – одиночные (рис. 38, 1) или колониальные бентосные прикрепленные организмы. Колонии массивные, ветвистые (рис. 38; 3, 4). Произошли от табулят в ордовике, расцвет подкласса приходится на силур-девон, окончательно вымерли в пермском периоде. Имеют карбонатный скелет. На начальной стадии развития в кораллите выделяются 4 вертикальные перегородки – септы, разделяющие полость коралла на 4 части. По мере роста появляются другие большие септы, а между ними развиваются малые – септы второго порядка (рис. 38; 1, 2а). В карбоне у некоторых ругоз в центре кораллита развился столбик (рис. 38, 1). Кроме вертикальных перегородок в кораллитах ругоз имеются еще и горизонтальные элементы – днища и пузыреобразные пластинки диссепименты (рис. 38, 2б). Ругозы имеют значение для стратиграфии большей части палеозойских отложений, и являются основными рифообразователями палеозоя. В карбоне они дали начало шестилучевым кораллам.
 Подкласс шестилучевые кораллы (Hexacorallia) – и скопаемые и ныне живущие, морские колониальные и одиночные организмы (рис. 33, 34), ведущие бентосный прикрепленный образ жизни. Скелет карбонатный, разделен септами на 6 частей. Имеют шестилучевую симметрию тела, количество щупалец кратное шести и септы трех порядков (рис. 36). Гексакораллы произошли от ругоз в каменноугольном периоде. В настоящее время они составляют основу современных рифов.
Подкласс шестилучевые кораллы (Hexacorallia) – и скопаемые и ныне живущие, морские колониальные и одиночные организмы (рис. 33, 34), ведущие бентосный прикрепленный образ жизни. Скелет карбонатный, разделен септами на 6 частей. Имеют шестилучевую симметрию тела, количество щупалец кратное шести и септы трех порядков (рис. 36). Гексакораллы произошли от ругоз в каменноугольном периоде. В настоящее время они составляют основу современных рифов.
Рис. 36. Ископаемый гексакоралл

Рис. 37. Подкласс Табулятоморфы:
1 – общий вид массивной колонии фавозитид,2 а – ячейка отдельного кораллита (видны поры), 2 б – общий вид массивной колонии михеленид, 3 – цепочечная колония хализитид (вид сверху), 3 а – увеличенные ячейки хализитид (поперечное сечение) с септальными шипиками

 4
4
Рис. 38. Подкласс Ругозы:
1 – общий вид скелета одиночного коралла, 2 а - поперечное сечение одиночного коралла (видны септы), 2 б – продольное сечение одиночного коралла (видны днища и пузырчатая ткань), 3 – общий вид массивной колонии, 4 – общий вид ветвистой колонии
 Подкласс Восьмилучевые кораллы (Octocorallia ) или альционарии — подкласс морских преимущественно колониальных организмов, с восемью щупальцами, насчитывающих около 2800 видов, которые разделяются на три отряда: альционарии (мягкие кораллы), роговые кораллы (в том числе благородный коралл, используемый как поделочный материал) и морские перья (рис. 39). Восьмилучевые кораллы распространены в основном в тропических морях, однако некоторые виды морских перьев населяют арктические воды.
Подкласс Восьмилучевые кораллы (Octocorallia ) или альционарии — подкласс морских преимущественно колониальных организмов, с восемью щупальцами, насчитывающих около 2800 видов, которые разделяются на три отряда: альционарии (мягкие кораллы), роговые кораллы (в том числе благородный коралл, используемый как поделочный материал) и морские перья (рис. 39). Восьмилучевые кораллы распространены в основном в тропических морях, однако некоторые виды морских перьев населяют арктические воды.
Рис. 39. Современный восьмилучевой коралл
Отдельные виды октокораллов могут существовать на больших глубинах. Обитающие на рифах представители Octocorallia обычно более терпимы к экстремальным значениям факторов среды, чем Hexacorallia, и заселяют участки недоступные для них. Скелет их карбонатный, развивается в мезоглее. Многие восьмилучевыекораллы используютхимические средства защиты, отравляяили отпугивая с их помощью потенциальных хищников. Это отличает их от шестилучевых, которые применяют для этой целистрекательные клетки.
Раздел Двусторонне-симметричные (Bilateria)
Двусторонне-симметричные организмы характеризуются способностью к активному перемещению. Они подразделяются на первично- и вторичноротых. Первичноротые получили название по основному признаку: ротовое отверстие взрослой особи находится там же, где и у личинки.
Подраздел Первичноротые (Protostomia)
К первичноротым относятся приапулиды (низшие черви), высшие (кольчатые) черви, членистоногие, моллюски, а также, возможно, мшанки и брахиоподы (Приложения 5, 6). Для палеонтологии важны и будут здесь рассмотрены высшие (кольчатые) черви (аннелиды).
Тип Кольчатые черви (Annelida), обитающие в водной и наземной среде, имеют пищеварительную, кровеносную, половую, нервную, мышечную и иногда дыхательную системы. Водные аннелиды обычно ведут неподвижный образ жизни, строя трубки из хитинового и известкового материала. Трубка может быть цилиндрической, прямой, изогнутой (рис. 40, 2, 3). Наружное отверстие трубки может закрываться крышечкой (рис. 40, 4). В результате полового размножения возникает плавающая личинка, которая потом садится на дно и дает начало кольчатым червям (рис. 40, 1). При массовом скоплении рода Serpula и близких ему родов образуются карбонатные породы – серпулиты. Наиболее древние аннелиды, вероятно, появились в венде. В вендских отложениях часто встречаются следы ползания, проедания грунта и зарывания в него, образованные различными организмами, в том числе, аннелидами. Достоверно доказанные аннелиды известны с кембрия. В современных морях аннелиды живут на различных глубинах, вплоть до ультраабиссали. За счет эволюции аннелид произошли членистоногие и моллюски.

Рис. 40. Кольчатые черви (Annelida): 1 – личинка современных аннелид,
2 – отпечаток кольчатого червя на сланце, 3 - общий вид колонии современных аннелид, 4 - крышечки раковин вымерших палеогеновых аннелид
Представители типа Членистоногие (Arthropoda), характеризуются сегментарным строением тела, и в настоящее время населяют все среды обитания органического мира Земли. В современной фауне они представлены 3 млн. видов, из которых около 1 млн. составляют насекомые. Тело артропод защищено хитиновым скелетом (панцирем), иногда пропитанным углекислым кальцием или фосфатом кальция. Рост особи осуществлялся в промежутках между обновлениями панциря (линьками). По строению головного отдела и органов дыхания членистоногие делятся на 4 подтипа: Трилобитообразные, Ракообразные (Жабродышащие), Хелицеровые и Трахейные. Древнейшие достоверно доказанные артроподы известны с венда.
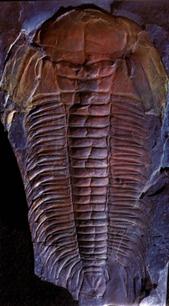
В подтипе Трилобитообразных (Trilobitomorpha) наибольшее значение для стратиграфии имеет класс Трилобиты(Trilobita) – вымершие морские животные (подвижный бентос), широко распространенные в отложениях нижнего и среднего палеозоя (более 10000 видов, принадлежащих к 2000 родов). Фактически трилобиты являются прародителями всех современных членистоногих. Они являлись первым подвижным бентосом на Земле: ползали по дну, зарывались в ил; некоторые плавали в придонных частях бассейнов. В ископаемом состоянии обычно встречаются фрагменты панциря и сравнительно редко – остатки конечностей, глаз, пищеварительного тракта. Скелет, в виде панциря, защищал лишь спинную часть тела трилобитов, а брюшная прикрывалась только кожистым покровом (рис.41, а,б). Скелет разделялся на сегменты: продольными бороздками обособлялась осевая часть тела и две боковые, а поперечными – головной и хвостовой щиты, туловищный отдел. В процессе эволюции у трилобитов выработалась способность сворачиваться, прижимая головной щит к хвостовому, в результате чего уязвимое брюшко оказывалось защищенным от хищников.
Головной щит, образованный 5-7 слившимися сегментами, подразделяется на осевую часть (глабель) и боковые (щёки), краевые части которых были подвижными. На щите располагались простые или фасеточные глаза. Туловище состояло из сегментов, число которых сильно варьировало (от 2 до 44). Каждый сегмент был снабжен парой двуветвистых конечностей, служивших для плавания или ползания по дну; их наружные ветви выполняли функцию жабр. Хвостовой щит, также как и головной, был образован сросшимися сегментами (от 1 до 30). По количеству сегментов и размерам трилобиты подразделяются на два подкласса: Малочленистые (Миомеры) и Многочленистые (Полимеры).
К миомерам относят мелких трилобитов (до 2 см), состоящих из 2-3 сегментов. Глаза обычно отсутствовали. Время существования – €-O. Полимеры характеризовались значительными размерами (1-70 см), а число сегментов у них было более 5. Они делятся на 7 отрядов. В процессе эволюции полимеров уменьшалась сегментация головного и хвостового щитов, сблизились их размеры (первоначально головной щит был значительно больше хвостового). Полимеры были распространены в течение всего палеозоя (рис.41, в), но уже в каменноугольных отложениях встречаются редко.
Класс Ракообразные(Crustacea) являются единственным классом подтипа, имеющего то же название – Ракообразные. Это – водные животные, тело которых подразделяется на голову, грудь и брюшко. На голове располагаются две пара усиков, играющих роль антенн, челюсти и глаза. К грудному отделу принадлежат конечности, служащие для движения, дыхания и захвата пищи. Для стратиграфии наибольший интерес представляют две группы примитивных ракообразных: Остракоды и Филлоподы.
Подкласс Остракоды – ракушковые рачки, населяющие морские, лагунные и пресноводные водоемы, подразделяются на 11 палеозойских и 5 мезо-кайнозойских отрядов. Размеры остракод варьируют в пределах 0,2-7 мм, но представители некоторых групп достигают 1-3 см. Тело особи заключено в двустворчатую раковину, построенную из органического вещества, пропитанного углекислым кальцием, (рис. 42, а, б). Рост тела происходил в промежутках между линьками, поэтому знаков роста на раковинах нет. Остракоды известны с кембрийского периода; пресноводные группы впервые появились в каменноугольных отложениях.
Из подтипа Хелицеровых класс Меростомовые (Merostomata) имеет наибольшую стратиграфическую значимость. Он разделяется на подклассы Мечехвостые и Эвриптероидеи (ракоскорпионы) (рис 43). Размеры эвриптероидей обычно составляли 10-20 см, но некоторые роды достигали 2 м в длину. С ордовика до перми они населяли солоноватые и пресные водоемы, являясь хищниками. Считается, что именно эти членистоногие стали первыми обитателями суши.
Класс Насекомых(Insecta) представлен наибольшим количеством видов (около 1 млн.). Тело насекомого длиной от 0,2 мм до 33 см состоит из головы, груди и брюшка, покрытых хитиновой оболочкой. Голова, образованная слиянием 4 сегментов, снабжена глазами, парой конечностей, преобразованных в антенны и тремя парами конечностей,

А б в
Рис. 41. Класс Трилобиты (Trilobita), подкласс Полимеры:
а – спинной отдел трилобита, защищенный панцирем, б – брюшной отдел трилобита, в - отпечаток кембрийского трилобита рода Paradoxides


А
Б
Рис. 42. Подкласс Остракоды (Ostracoda): а –схема внутреннего строения остракод (видны конечности, глаз и жабры), б – современная остракода (увелич.)

Рис. 43. Отпечаток ракоскорпиона Eurypterus remipes на песчанике
превращенных в челюсти. Грудь состоит из трех сегментов, каждый из которых снабжен парой ног. Кроме того, второй и третий сегменты груди несут крылья. Брюшко состоит из 5-11 сегментов, не имеющих конечностей. Насекомые появились в девоне, а в настоящее время переживают эпоху расцвета. Важны для стратиграфии, начиная с позднего палеозоя. Насекомые играли важную роль в эволюции растительности, особенно цветковых. Из беспозвоночных это единственная группа, освоившая полет в воздушном океане.
Тип Моллюски (Molluska) подразделяется на 10 классов, из которых наибольшее значение для стратиграфии и фациального анализа имеют Двустворчатые, Брюхоногие и Головоногие моллюски, а также Тентакулиты (Приложения 6 и 7). Тело моллюска, как правило, имеет двусторонне-симметричное строение, подразделяясь на голову, туловище и ногу. Обычно оно защищено известковой раковиной и только у некоторых групп она отсутствует. Характерной особенностью моллюсков является обитание в разных средах, что обусловило высокую изменчивость морфологических признаков. Особым разнообразием характеризуются нога и раковины. По видовому разнообразию (150 тыс. видов) моллюски стоят непосредственно за членистоногими. Моллюски известны, начиная с кембрия.
Представители класса Двустворчатые моллюски(Bivalvia) – морские или пресноводные животные, тело которых заключено в двустворчатую раковину, причем одна створка является зеркальным отражением другой (рис. 44, б). Мягкое тело не имеет обособленной головы, но есть мускульная нога для прикрепления. Прикрывающая тело мантия – особая складка кожи, выделяющая перламутр, вещество, которое является основой для жемчуга (рис. 44, г). Жемчуг – это органогенная конкреция, растущая внутри раковины вместе с моллюском.
Раковины двустворок имеют выступающую макушку и край с замочным аппаратом, скрепляющим их друг с другом. Сочленение створок осуществлялось с помощью связки и замка, состоящего из выступов (зубов) на одной створке и зубных ямок – на другой (рис. 44, а). Размеры раковин варьируют в широких пределах от долей миллиметра и почти до метра. Наиболее крупные характерны для приэкваториальных зон. Форма раковин разнообразна до причудливости: округлые, эллиптические, конические, рогообразные, шиповатые (рис. 44,) и т.д. Систематика современных и ископаемых двустворок строится по-разному: современные классифицируются по

А б


В г
Рис. 44. Двустворчатые моллюски: а – внутренняя часть створки с зубным аппаратом, б – раковина современного рода Cardium, в - наружная часть створки с шиповато-ребристой скульптурой, г - современная двустворка с жемчужиной
 строению мягкого тела, ископаемые – по строению замка. По этому принципу выделяется шесть отрядов, различающихся наличием или
строению мягкого тела, ископаемые – по строению замка. По этому принципу выделяется шесть отрядов, различающихся наличием или
Из зон Мирового океана двустворки предпочитают сублитораль. Одни группы медленно передвигались по поверхности грунта, другие переходили к неподвижному образу жизни, прирастая к субстрату. Некоторые двустворки за счет ритмичного открывания и захлопывания створок освоили перемещение в придонном слое воды, другие зарывались в илистые грунты или сверлили камни или дерево. Нередко образовывали массовые скопления – банки. Некоторые двустворки (устрицы, гребешки) являются продуктами питания человека.
Двустворки появились в кембрии, но настоящего расцвета достигли только в мезозое и широко распространены до наших дней. Во второй половине мезозоя в теплых бассейнах появились массивные прикрепленные двустворки (рудисты), конвергентно сходные с ругозами. Они активно участвовали в рифостроении. Бивальвии имеют значение для фациального анализа, а также для стратиграфического расчленения мелового, палеогенового и неогенового периодов.
Класс Брюхоногие моллюски (Gastropoda) включает современные и вымершие виды (рис. 45 – 48) и является наиболее многочисленным классом типа моллюсков, освоившим разнообразные среды обитания (море, пресные водоемы, суша). Они характеризуются асимметричным телом с обособленной головой, на которой располагаются органы зрения, обоняния и осязания в виде антенн. Тело заключено в раковину, спирально закрученную в горизонтальной или вертикальной плоскости (рис. 45, 46). Большинство гастропод ведет подвижный образ жизни, передвигаясь с помощью ноги c присоской, помещенной на брюшной полости. В зависимости от строения органов дыхания выделяются подклассы Переднежаберные, Заднежаберные и Легочные.
Подкласс Переднежаберные характеризуется расположением жабер впереди сердца. Это – преимущественно морские организмы, обладающие спиральнозавитой (рис.47, а) или колпачковидной (рис. 47, б) раковиной. Имеется нога в виде подошвы или киля. Переднежаберные появились в кембрии и существуют до настоящего времени. По традиции выделяются отряды Археогастроподы, Мезогастроподы и Неогастроподы, хотя сейчас принято более дробное деление.
Подкласс Заднежаберные обладает одной жаброй, расположенной позади сердца. Раковина редуцирована и нередко отсутствует. Преимущественно морские организмы, населяющие все зоны Мирового океана. Некоторые группы освоили пресноводные водоемы. Заднежаберные известны с каменноугольного периода до настоящего времени. В качестве особого отряда Заднежаберных выделяются Крылоногие моллюски – Птероподы (P – ныне) – характеризующиеся мелкими раковинами (1-2 мм). Нога их изменена в два крыловидных плавника (рис.48). После смерти животных за счет скопления раковин образуются птероподовые илы.
Подкласс Легочные (Pulmonata), (рис. 45) существующие с каменноугольного периода, освоили субаэральную среду, в связи с чем, жабры у них заменились легкими. В составе подкласса имеются раковинные и безраковинные формы. Большинство легочных моллюсков являются растительноядными, некоторые – хищниками или паразитами. Гастроподы используются для стратиграфии кайнозойских отложений. В последние годы предпринимаются попытки использования для стратиграфии палеозойских и мезозойских представителей этого класса.


|
Раковина вертикально закрученная, с крышечкой
на задней части тела. Голова обособлена и
снабжена органами зрения, обоняния и осязания
А б
Рис. 47. Представители подкласса Переднежаберные:
а - хищные гастроподы рода Conus (палеоген-ныне),
б – различные виды современных гастропод рода Patella (мел-ныне)

Рис. 48. Подкласс Заднежаберные: современные птероподы
Класс Головоногие моллюски (Cephalopoda) является наиболее важным для стратиграфии фанерозоя классом типа Моллюски. У большинства цефалопод тело заключено в прямую или спирально закрученную многокамерную раковину. Строение межкамерной перегородки и её отпечаток на раковине – перегородочная линия – являются важными классификационными признаками. Эволюция цефалопод сопровождалась усложнением очертаний перегородочной линии. Тело располагается в последней камере, а остальные заполнены газом, благодаря чему увеличивается плавучесть раковины. Соединение камер обеспечивается сифонной трубкой (рис. 49, б). Частично заполняя камеры водой, особь могла регулировать свое положение в толще воды. Нога цефалопод преобразована в венчик щупалец и воронку, выбрасывающую воду, что обеспечивает реактивное движение особи. В последние годы несколько изменена классификация цефалопод. Они разделены на 7 подклассов, из которых два (наутилоидеи и колеоидеи) представлены в современной фауне, а остальные – вымерли. У шести подклассов тело заключено в раковину, а у седьмого (колеоидеи) – раковина находится внутри тела, выполняя роль внутреннего скелета.
Подкласс Наутилоидеи, известный в ископаемом состоянии с позднего кембрия, в современной фауне представлен единственным родом Nautilus (отряд Наутилиды). Тело наутилоидей было заключено в прямую или спирально закрученную раковину, межкамерная перегородка прямая (рис.49, а, б).


А б
Рис. 49. Род Nautilus: а – современный наутилус
б – ископаемый наутилус с прямой перегородочной линией
Подкласс Ортоцератоидеи (O –T) характеризовался прямой раковиной, достигавшей в длину 1-1,5 м. Межкамерная перегородка линзовидно-изогнутая, перегородочная линия прямая или почти прямая. Сифон субцентральный (рис. 50). К подклассу Эндоцератоидеи(O) принадлежали наиболее крупные цефалоподы, достигавшие в длину 3-4, а в отдельных случаях до 10 м. Поперечное сечение – круглое или овальное. На сифон, расположенный на брюшной стороне и занимающий краевое положение, могло проходиться до 1/3 поперечного сечения (рис. 50). Прямая перегородочная линия. Подкласс Бактритоидеи являлся небольшой по числу видов группой, обладавшей прямой раковиной. Узкий сифон занимал краевое положение. Подкласс рассматривается как возможный предок аммоноидей.

Рис. 50. Реконструкция ордовикского моря: 1 – плавающие ортоцератоидеи,
2 – охотящийся на трилобитов, представитель эндоцератоидей
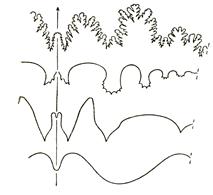
Подкласс Аммоноидеи, характеризующийся наибольшим видовым разнообразием, появился, по-видимому, в девонском периоде и вымер в конце мелового (рис. 53). Аммоноидеи со спирально закрученной раковиной существовали с девона до мела и фигурируют в качестве мономорфных; с прямой, коленообразно изогнутой или крючковатой раковиной – как гетероморфные. Они появились в позднем триасе, а вторично возникли в средней юре, но наиболее широкое распространение получили во второй половине мела. Жилая камера у разных представителей этого подкласса занимала от половины до 2 оборотов раковины. В ней отсутствуют межкамерные перегородки, а в остальных камерах они имеют сложноскладчатый характер. Перегородочная линия состоит из лопастей и сёдел. Изгибы перегородочной (лопастной) линии в зависимости от направления изгиба (к жилой камере или к заднему краю раковины) называются либо седлами, либо лопастями. У аммоноидей наблюдается 4 типа перегородочной линии: агониатитовая (рис. 52, а), гониатитовая (рис. 52, б), цератитовая (рис. 52, в) и аммонитовая (рис. 52, г). Агониатитовая линия характеризуется округлым характером нерасчлененных сёдел и лопастей. При гониатитовой линии сёдла и угловатые лопасти не расчленены. При цератитовой – расчленены все лопасти, при аммонитовой – расчленены и сёдла, и лопасти. Строение перегородочной линии, её усложнение во времени, а также положение сифона лежат в основе расчленения подкласса Аммоноидеи на восемь стратиграфически значимых отрядов.
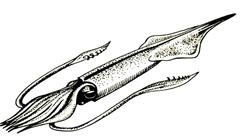
Подкласс Колеоидеи объединяет группы головоногих моллюсков, у которых раковина играла роль внутреннего скелета. Это – активно плавающие хищники (рис. 51, а). Древние колеоидеи объединены в надотряд Белемноидеи (C – K). В ископаемом состоянии обычно сохраняется основная часть внутреннего скелета – ростр – массивное, заостренное на конце цилиндрическое образование, состоящее из арагонита (рис. 51, б). В процессе онтогенеза форма ростра нередко менялась. В углублении расширенного конца ростра – альвеоле – располагались важнейшие части тела особи. Современные колеоидеи – надотряд Декабрахии (J – Q) – представлены осьминогами, каракатицами, кальмарами. К ним, в частности, принадлежат наиболее крупные представители беспозвоночных – гигантские кальмары, достигающие в длину 18 м. Представители третьего надотряда – Октобрахии (K – Q) не имеют скелета, в силу чего их стратиграфическая значимость невелика.

|
|

,
Рис.51. Надотряд белемноидеи: а –реконструкция древних форм
б – ростры ископаемых белемноидей (J-K
 г
г
В
Б
Рис. 52. Перегородочные линии
Дата публикования: 2014-10-25; Прочитано: 3845 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
