 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Охотятся на аммонитов
|
|
В состав завроптеригий входят группы Плезиозавры и Плиозавры. Плезиозавры (T2 – K) обладали длинной шеей, маленькой головой, веретенообразным телом, достигавшим в длину 16 м. Плиозавры (J1 – K) имели большую голову с короткой шеей, бочковидное или веретенообразное тело длиной до 15 м. Плакодонты (средний триас – ранняя юра), по облику напоминавшие черепах, питались двустворчатыми моллюсками и донными водорослями. Передние зубы были превращены в резцы, задние – приспособлены для раздавливания пищи. Нередко зубной аппарат заменялся клювом.
Подкласс Лепидозавры («чешуйчатые ящеры») характеризовались особой формой черепа с двумя височными окнами и нередко наличием отверстия для «теменного глаза». Тело лепидозавров было покрыто чешуйками или пластинами. По образу жизни лепидозавры преимущественно наземные животные, реже – водные. Некоторые представители лепидозавров лазили по деревьям и скалам, другие – обладали способностью к планированию. Подразделяются на отряды Эозухии, Клювоголовые и Чешуйчатые. Эозухии (P – T1), напоминавшие ящериц и крокодилов, отличались от них строением зубного аппарата. Длина их тела не превышала 0,5 м. Клювоголовые (мезозой – ныне) в современной фауне представлены единственным видом – гаттерией. К чешуйчатым принадлежали крокодилоподобные мозазавры (поздний мел), ящерицы (T – ныне) и змеи ( K – ныне). У мозазавров размеры тела достигали 15 м, узмей – 11 м, у ящериц – 4 м.
Подкласс Архозавры – наиболее разнообразный подкласс рептилий. Они различаются по облику, размерам, строению скелета и образу жизни. Общими признаками архозавров являются тип черепа, имеющий два височных окна и зубы, формирующиеся в отдельных ячейках, но не имеющие корней. Подкласс разделен на надотряды Текодонты, Динозавры и Птерозавры, но это разделение, по-видимому, является условным. В частности, птерозавры, в значительной мере приспособленные к полету, целесообразно выделить в особый подкласс.
Надотряд Текодонты (P3 – T), являющиеся наиболее древней группой, вероятно, произошли от эозухий, от которых отличаются отсутствием отверстия для «теменного глаза» и нёбных зубов. По образу жизни текодонты напоминали динозавров. Максимальные размеры тела достигали 6 м. Подразделяются на плотоядных псевдозухий и растительноядных фитозавров.
Надотряд Динозавры – наиболее распространенные мезозойские рептилии. Они появились в триасовом периоде, характеризовались широчайшим разнообразием в юрском и меловом, а затем вымерли в конце мела. Причины исчезновения динозавров являются одной из дискуссионных проблем палеонтологии. Яйца динозавров достигали 30 см. До недавнего времени палеонтологи подчеркивали низкий уровень психической деятельности динозавров. Однако исследования последних лет показали, что это не совсем так. Было установлено, что динозавры жили колониями и по уровню заботы о потомстве не уступали птицам. Некоторые группы динозавров совершали значительные переходы в меридиональном направлении, напоминавшие сезонные перелеты птиц. В настоящее время большинство исследователей подразделяет динозавров на отряды Ящеротазовых и Птицетазовых.
Ящеротазовые динозавры (T2 – K) были двуногими и четвероногими формами, хищными и растительноядными, наземными и прибрежно-полуводными (рис. 71 А, Б). Размеры их весьма различны: от 0,2 до 35 м. Хищные ящеротазовые динозавры – звероногие – были двуногими. К ним относятся страусоподобные целурозавры (средний триас – мел) и карнозавры (юра – мел). Среди карнозавров встречались такие гиганты как тираннозавры, достигавшие в высоту 4-5 м, а в длину – 10-15 м. Размер черепа составлял 1,5 м. Огромная пасть несла краевой ряд конических зубов разного размера. Растительноядные ящеротазовые динозавры – ящероногие – (поздний триас – мел) были четвероногими формами. Маленькая голова располагалась на длинной шее. Хвост мог превышать длину тела. Одной
из крупнейших форм ящероногих был диплодок, достигавший в длину 25-35 м при высоте 4-6 м. По-видимому, значительную часть времени он проводил в воде, уменьшая, таким образом, свой вес (рис.71 А).
Птицетазовые динозавры характеризовались меньшим разнообразием, по сравнению с ящеротазовыми. Они были растительноядными наземными животными четвероногими или двуногими.
Среди четвероногих птицетазовых выделялись стегозавры, анкилозавры и цератопсы. Стегозавры (J –K1) имели небольшую голову и хвост с шипами. На спине располагался ряд пластин высотой до 1 м неясной функции. Размеры тела достигали 6-10 м. Анкилозавры (J2 – K) имели толстые скульптированные панцирные пластины с шипами, что делало их похожими на парейазавров. Цератопсы – рогатые ящеры (поздний мел) – имели снабженную рогами голову, занимающую до 1/3 туловища. Высота их достигала 3 м, длина – 6-8 м. Зубы многорядные, передние челюсти нередко клювовидные.
Из двуногих птицетазовых динозавров наиболее известны игуанодоны и утконосые динозавры (гадрозавры). Игуанодоны (ранний мел) имели мощные трехпалые задние конечности, выполнявшие опорные функции. Передние пятипалые конечности были укорочены. Лопатовидные зубы располагались в несколько рядов. Гадозавры (поздний мел) имели костный гребень на голове, широкий утинообразный клюв, мощные задние ноги и укороченные передние, длинный хвост. Высота животных достигала 5-15 м. Зубы располагались кассетами в несколько рядов. Их количество достигало тысячи. Гадозавры вели прибрежно-полуводный образ жизни, обитая вблизи водоемов.
В современной фауне представлена лишь одна группа архозавров – отряд (?) Крокодилы, в значительной мере вымерший. Среди современных рептилий крокодилы выделяются наиболее высоким

Рис. 71 А. Ящеротазовые. Диплодоки



|
уровнем развития (наличие четырехкамерного сердца, развитых легких, вторичного нёба).
Надотряд Птерозавры, (рис.72) характеризовавшиеся способностью к полетам, имели крыловидную кожистую перепонку между туловищем и пятым пальцем передних конечностей. Кости скелета были полыми. По размерам и строению тела птерозавры подразделялись на отряды Рамфоринхи и Птеродактили. Рамфоринхи (поздний триас – юра) были небольших размеров (до 0,5 м; в размахе крыльев – до 2,5 м). Они имели длинный хвост, пятипалые задние конечности, короткую шею, хорошо развитые зубы. Птеродактили (J – K) имели короткий хвост, четырех-пятипалые задние конечности, укороченную шею, слабо развитый зубной аппарат или беззубый клюв. Размеры особей были различны: от 30-40 см до гигантов, достигавших в размахе крыльев 10-12 м. Многие из них обладали способностью к парящему полету. Птеродактили обитали вдоль побережий морей и внутриконтинентальных водоемов, питаясь рыбой.
Подкласс Зверообразные (C3 – J2) были четвероногими рептилиями, считающимися предками млекопитающих. Большая их часть вела наземный образ жизни, реже – полуводный. По типу питания принадлежали к хищникам; растительноядные формы встречались сравнительно редко. Древнейший отряд – пеликозавры (C3 – P) – отличался от млекопитающих строением нижней челюсти, характером сочленения с черепом позвоночного столба, степенью дифференциации зубного аппарата. Размеры пеликозавров не превышали 2-3 м. Отряд Терапсиды (P3 – J2) характеризовался лучшей дифференциацией зубного аппарата. Они подразделялись на четыре группы, различавшиеся размерами, строением зубов, образом жизни. К зверозубым принадлежали хищные иностранцевии (P3), достигавшие в длину 3-3,5 м. Наиболее близки к млекопитающим были цинодонты (собакозубые ), размером с волка. В триасе они захватили всю сушу и вымерли в средней юре последними из терапсид (рис.73).
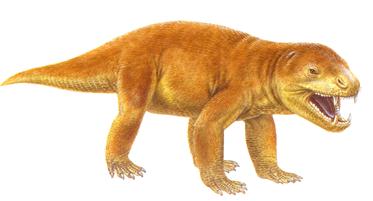
Рис.73. Цинодонты ( внешнепохожие на собак)
Класс Птицы(Aves) – наиболее многочисленный класс наземных позвоночных – произошли от небольших мезозойских рептилий, передвигавшихся на двух ногах. По одному предположению, предками птиц были быстро бегающие и периодически взлетавшие наземные рептилии открытых пространств, по другому – древесные пресмыкающиеся, перелетавшие от дерева к дереву. Ключевым для птиц является набор признаков: наличие перьев, преобразованных из роговых чешуй; мощная грудина и задний крестец, образовавшиеся из сросшихся позвонков; облегченные, частично полые кости; интенсивный обмен веществ, обеспечивающий теплокровность; передвижение на утолщенных задних конечностях; размножение яйцами. Некоторые из этих признаков присущи и другим классам, но их совокупность характерна лишь для птиц. Размер птиц – от 3 см до 3 м. В размахе крыльев альбатросовые достигают 5 м (Приложение 7).
В эволюции птиц до недавнего времени выделялось два подкласса: Ящерохвостые и Веерохвостые. Типичным представителем ящерохвостых считался археоптерикс (рис.74), семь экземпляров которого было найдено в верхнеюрских отложениях Баварии (ФРГ). Археоптерикс обладал длинным хвостом из многочисленных позвонков, вдоль которых располагались перья. Зубы археоптерикса были текодонтного типа, роговой чехол клюва отсутствовал. В 1987 году в верхнетриасовых отложениях Техаса (США) было найдено 3 экземпляра ранее неизвестного птицеподобного существа, названного протоависом. Несмотря на более древний возраст, протоавис имел большее сходство с веерохвостыми птицами, чем археоптерикс. Исходя из этой находки, была выделена третья наиболее древняя группа птиц (особый подкласс), являющаяся прямым предком веерохвостых. С этих позиций ящерохвостые (и их характерный представитель археоптерикс), оказываются тупиковой ветвью эволюции класса птиц.
Веерохвостые птицы (K – ныне) имеют короткий хвостовой отдел позвоночника, состоящий из двояковогнутых или седловидных позвонков, к которым веерообразно прикреплены хвостовые перья. По строению клюва веерохвостые птицы делятся на зубастых (K3) и беззубых (современных), имеющих на клюве роговой чехол. Среди птиц известны плотоядные, растительноядные и всеядные формы. Подавляющее большинство птиц способно к полету. Несравненно меньшее распространение имеют бегающие или водоплавающие группы (страусы, пингвины). Полные скелеты ископаемых птиц встречаются крайне редко. Иногда сохраняются перья, скорлупа яиц, следы птиц. Большинство находок происходит из озерных, болотных и морских отложений. а б
Рис. 74. Археоптерикс: а – реконструкция, б – отпечаток на сланце (J)
Класс Млекопитающие (Mammalia), вскармливающие детенышей молоком, резко различны по размерам, облику, образу жизни (Приложение 10). Масса их тела варьирует в широчайших пределах: от землеройки весом в несколько грамм до китов весом в десятки тонн. Характерной особенностью млекопитающих является теплокровность и защитные приспособления, обеспечивающие постоянство температуры тела (волосяной покров или мощная жировая прослойка). Зубы дифференцированы. Разделение млекопитающих на подклассы базируется на способе размножения.
Подкласс Первозвери (T3 – ныне) вскармливают детенышей молоком, но размножаются яйцами. Строение скелета и наличие клоаки, куда открываются пищеварительный тракт, мочевые и половые протоки, сближают их с пресмыкающимися. В современной фауне подкласс представлен отрядом Однопроходные, достоверно известным с неогена. Представители этого отряда – утконос и ехидна – в настоящее время населяют Австралию и Новую Гвинею. Это – небольшие животные (длина тела до 60 см), ведущие наземный и полуводный образ жизни. К вымершим представителям этого подкласса принадлежат отряды Многобугорчатые (T3 – P2) и Докодонты (J3 – K1).

Рис. 75. Морганукодон – одно из самых древних млекопитающих планеты (T)
Условно в данный подкласс (рис.75) включают небольших крысоподобных животных, принадлежащих к отряду Трикодонты. Классификация данного подкласса остается дискуссионной.
Подкласс Сумчатые (поздний триас? – поздний мел – ныне) рождают недоразвитых детенышей, которые вскармливаются молоком и донашиваются самкой в особых кожаных сумках. Выводковая сумка поддерживается костями таза. Степень дифференциации зубов и их число различно у разных групп. Из сумчатых наиболее известны кенгуру, опоссумы и коалы. Кроме того, в неформальной номенклатуре часто фигурируют формы, конвергентно сходные с соответствующими группами плацентарных (сумчатые кроты, мыши, куницы, волки). Размеры сумчатых варьируют от нескольких сантиметров до 3 м (кенгуру). Сумчатые представлены в современной фауне трех континентов: Австралии (7 отрядов), Северной и Южной Америк (3 отряда). Древние сумчатые были известны на всех континентах. Предполагается наличие общего предка сумчатых и плацентарных млекопитающих.
Подкласс Плацентарные ( Высшие млекопитающие) появились в позднем мелу, а господствующее положение заняли с палеогена (Приложение 10). Мел-палеогеновый рубеж рассматривается как начало господства высших (плацентарных) млекопитающих, являющихся наиболее высокоорганизованными животными. Характерной особенностью этой группы является наличие плаценты – особого органа, осуществляющего связь материнского организма с детенышем до момента его рождения. Другими важнейшими особенностями плацентарных являются сложное строение головного мозга, отсутствие выводковой сумки, рождение беззубых детенышей, способных сосать материнское молоко.
Систематика плацентарных основана на строении зубного аппарата, черепа и конечностей. Выделяется около 30 отрядов, из которых в современной фауне представлено около 20. В раннемеловую эпоху появились насекомоядные, зайцеобразные и приматы, в позднемеловую – древние копытные. В раннепалеогеновых отложениях известны рукокрылые, хищники (креодонты и настоящие хищники), грызуны, непарнокопытные; в среднепалеогеновых – хоботные, китообразные, парнокопытные. Широчайшая радиация, обусловившая наступление «века млекопитающих», начинается с эоцена.
Отряд Насекомоядные (K1 – ныне)– всеядные животные с длиной тела 3,5-22 см. Зубной аппарат дифференцирован, но не специализирован. Мозг маленький, большие полушария гладкие.
Отряд Рукокрылые (P1 – ныне) представлен подотрядами Летучие мыши и Крыланы, различающимися, главным образом, размерами (первые – 2,5-14 см, вторые – до 40 см, в размахе крыльев – до 1,7 м). Один конец летательной кожистой перепонки рукокрылых крепится на пальцах передней конечности, другой – на задних конечностях. В зубном аппарате хорошо развиты клыки; предкоренные и коренные зубы слабо дифференцированы. Большие полушария головного мозга имеют малое количество извилин. По способу питания рукокрылые принадлежат к насекомоядным, растительноядным и кровососущим (вампиры).
Отряд Древние хищники (креодонты), включающий только ископаемые формы, существовал в течение палеогена и раннего неогена. Размеры тела варьировали от 10 см до 2,5 м. Головной мозг имел небольшие размеры, большие полушария были почти гладкими. Короткие конечности были преимущественно пятипалыми. Зубы – бугорчатые или бугорчато-режущие. Креодонты произошли от вымершей группы насекомоядных.
Отряд Настоящие хищники (Фиссипедии) имеют тело размером от 10 см до 4 м. Передние конечности обычно пятипалые, задние – четырехпалые. Характерны плотоядные зубы с режущими гребнями. Большие полушария головного мозга несут многочисленные извилины. Настоящие хищники подразделяются на собакоподобных (палеоген – ныне) и кошкоподобных (средний палеоген – ныне). Собакоподобные, преимущественно, вынюхивают добычу, кошкоподобные – высматривают. В разных филогенетических линиях кошек неоднократно появлялись саблезубые формы.
Отряд Зайцеобразные (K1 – ныне) включает в себя как ископаемые, так и современные группы. К последним принадлежат пищуховые и зайцевые, имеющие размеры тела 30-50 см. Передние конечности пятипалые, задние – четырехпалые. Верхняя челюсть несет постоянно растущие резцы. Клыки – отсутствуют, предкоренные и коренные зубы слабо дифференцированы. Полушария головного мозга почти гладкие, с малым количеством извилин. Ранее грызуны и зайцеобразные включались в один отряд, но позднее было доказано независимое от грызунов происхождение зайцеобразных от одной из групп насекомоядных.
Отряд Грызуны (P1 – ныне) представлен разнообразными группами с размерами тела 5-130 см (беличьи, хомяковые, мышиные и другие). Передние конечности пяти- или четырехпалые, задние – пяти- четырех- или трехпалые. Вторые резцы растут постоянно, клыки и предкоренные зубы отсутствуют. Число коренных зубов зависит от пищи. Полушарии почти гладкие, с малым количеством извилин. Грызуны имели общих предков с насекомоядными. Зубы грызунов, хорошо сохраняющиеся в ископаемом состоянии, используются для стратиграфии континентальных неоген-четвертичных отложений.
Копытные, по-видимому, являются филогенетически сборной группой, включающей отряды, характеризующие разные геологические периоды.
Отряд Кондилартры (K2) представлен животными с длинным хвостом, короткими пятипалыми конечностями, острыми копытами. Размер тела варьировал от 30 см до 2 м. Произошли от насекомоядных, но в палеогене в процессе радиации разделились на группы по способу питания. Одни стали родоначальниками китообразных и ластоногих, другие – предками непарнокопытных, хоботных, парнокопытных.
Отряд Непарнопалых (Непарнокопытных) объединяет несколько современных и ископаемых групп. Длина тела этих животных варьировала от 0,4 до 7 м; высота достигала 2-5 м. Конечности большинства непарнокопытных удлиненные, с различно оформленными копытами. Число пальцев на передних и задних конечностях – три или один, причем средний был наиболее крупным. В процессе эволюции число пальцев уменьшалось за счет редукции боковых.
К современным непарнокопытным принадлежат лошадиные (палеоген – ныне), тапировые (поздний палеоцен – ныне) и носороговые (средний палеоген – ныне); к ископаемым – гиракотериевые и титанотериевые (P2-3), халикотериевые (P2 – Q1), индрикотериевые (P3 – N1). Лошадиные вначале не превышали в высоту 1 м. Они обладали четырехпалыми передними и трехпалыми задними конечностями. Наибольшее распространение имели в Северной Америке, откуда неоднократно мигрировали на территорию Евразии. Носороговые характеризовались короткими массивными конечностями, вначале четырехпалыми, позднее – трехпалыми. Большинство носороговых были безрогими. Зубы характеризовались редукцией клыков и верхних резцов. Нижние резцы превратились в бивни. Места обитания носороговых весьма разнообразны. В раннечетвертичное время широкое распространение имел шерстистый носорог – предмет охоты древнего человека.
Из вымерших непарнокопытных наибольшими размерами характеризовались индрикотериевые. Их максимальные размеры достигали 7 м в длину и 5 м в высоту. Зубной аппарат был частично редуцирован. Передние резцы превратились в бивни, в верхней челюсти направленные вниз, а в нижней – вверх. Объем мозга был невелик, шея длинная. Конечности массивные трехпалые. Индрикотериевая фауна была широко распространена в течение олигоцена и миоцена в лесостепной зоне Евразии.
Специфическим признаком отряда Хоботные, является наличие хобота, образовавшегося за счет сращения носа и верхней губы. Конечности пятипалые, от коротких и средних до длинных и массивных. Отряд был широко распространен в течение неогена и плейстоцена, а в настоящее время почти вымер. Древние хоботные были высотой до 1 м, но в процессе эволюции достигли 4,5-5 м. Изменение размеров и облика связано с переходом от жизни в лесах к обитанию на открытых пространствах. Подразделяются на подотряды Меритерии, Дейнотерии и Слонообразные, из которых последние разделены на мастодонтов и слонов. Хоботные возникли в Африке, а затем распространились в Евразию и Северную Америку.
Отряд Китообразные – вторичноводные млекопитающие, постоянно живущие в водной среде. Дыхание легочное, но могут находиться под водой до 1,5 часов. По внешнему виду конвергентно сходны с рыбами. Роль плавников выполняют сильно укороченные пятипалые передние конечности. Задние конечности и кости таза сильно редуцированы, носовые отверстия смещены назад. Отряд подразделяется на группы Древние, Зубатые и Беззубые киты. Филогенетическое родство зубатых и беззубых китов вызывает сомнение.
Древние киты (эоцен – миоцен) по строению зубного аппарата напоминают кондилартр. К зубатым китам (эоцен – ныне) принадлежат дельфины, кашалоты, нарвалы, клюворылые. Зубы однообразны, число их увеличивается до 240 или уменьшается до двух. Зубатые киты – морские, реже речные или озерные животные, питающиеся рыбой, головоногими моллюсками, ракообразными. Длина тела 1,2 – 20 м. Беззубые (усатые) киты (олигоцен – ныне) вместо редуцированных зубов содержат в ротовой полости многочисленные роговые пластины («китовый ус»). Беззубые киты – морские животные, питающиеся планктоном или стайными мелкими рыбами, процеживая пищу через «усы». Длина тела до 33 м.
Отряд Парнокопытные подразделяется на жвачных и нежвачных. Современные представители жвачных появились в олигоцене (бычьи, оленьи), неогене (оленьковые, жирафовые, антилокапры) и плейстоцене (полорогие: газели, сайгаки, козы, бараны). Из нежвачных современных групп в эоцене появились свинообразные и пекари, неогене – гиппопотамы. Верблюдов и лам обычно выделяют из состава парнокопытных в особый отряд Мозоленогие. Длина тела современных парнокопытных варьирует в пределах 1,5-4,5 м, высота – 2-5,5 м. Кроме того, в составе данного отряда имеются ископаемые группы. Обособление парнокопытных в особую группу обусловлено их отличиями в способе передвижения и мало связано с особенностями строения зубного аппарата.
Отряд Приматы (Приложение 10) произошел в раннемеловую эпоху от группы животных близких к насекомоядным. Он подразделяется на подотряды Полуобезьяны (Низшие приматы) и Обезьяны (Высшие приматы). К первому подотряду (эоцен – ныне) принадлежат лемуры, долгопяты, лори и близкие им формы. Ко второму – относятся широконосые обезьяны Америки, подразделяющиеся на игрунковых и цепкохвостых, а также узконосые обезьяны Старого Света (мартышкообразные и человекообразные). Длина тела обезьян варьирует от 10 см до 1,5-2 м. Пятипалые конечности обладают высокой хватательной способностью. Останки древнейшего представителя высших приматов найдены в эоценовых отложениях Мьянмы (Бирма). В Африке останки высших приматов известны с олигоцена. Предок человека и человекообразных обезьян – дриопитек п оявился 25-30 млн. лет назад в олигоцене. Был жителем влажных тропических лесов. Вел древесный образ жизни, питаясь растениями и насекомыми (рис.76).

Рис. 76.
Дата публикования: 2014-10-25; Прочитано: 2083 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
