 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Условия возникновения циркуляции
|
|
1. Время между посылкой двух импульсов возбуждения (Т) должно быть меньше периода рефрактерности R2 >T.
2. Длина окружности кольца должна быть больше длины волны возбуждения.
Длина циркулирующей волны (при скорости 3 м/с и периоде примерно в 0,3 с) в путях проведения может быть около 1 м, поэтому ее появление анотамически невозможно. Однако в узлах медленного проведения возбуждения, где скорость может быть около 0,01 м/с, а длина волны порядка нескольких мм. Поэтому механизм циркуляции волны возбуждения в сердце возможен (например в области артиовентрикулярного узла и в зонах с замедленным проведением автоволн.
26. АВТОВОЛНОВЫЕ ПРОЦЕССЫ В СЕРДЕЧНОЙ МЫШЦЕ
Мышечная ткань представляет собой возбудимую среду. Возбуждение, сообщаемое сетью нервных волокон, вызывает механохимические процессы. Процессы в возбудимых средах имеют важнейшее значение в физиологии, в частности, в физиологии сердечной мышцы.
В возбудимой среде распространение импульса происходит без затухания за счет энергии клеток. Импульс генерируется каждой точкой среды. Пример однородной возбудимой среды – нервное волокно. Каждая точка возбудимой среды может находиться в одном из трех состояний: покоя, возбуждения, рефрактерности. Если на малый участок волокна, находящегося в состоянии покоя, подать возбуждение, то он перейдет в возбужденное состояние, в котором будет находиться некоторое время t, затем перейдет в рефрактерное состояние, в котором будет пребывать tR. В период рефрактерности tR участок не реагирует на возбуждающий сигнал, поэтому данный участок не может возбуждаться с частотой, большей t -1R.
В возбудимых средах реализуются автоволновые процессы. Источники волн могут возникать и в таких средах, в которых ни один из элементов не способен возбуждаться самопроизвольно, то есть не является автоколебательным. Возникновение таких источников обусловлено особыми свойствами среды и волны возбуждения.
При рассмотрении реакций Белоусова – Жеботинского было отмечено существование источников волн двух типов – с ведущими центрами (ЭХО), посылающими концентрические волны, и с ревербераторами – вращаю-щимися спиральными волнами. Возникновение и последующее размно-жение ревербераторов играет важнейшую роль в механизме фибриляции - очень опасной сердечной аритмии. Аритмия означает нарушение синх-ронизации распространения волн в сердечной мышце: вместо синхронных сокращений возникают беспорядочные колыхания, и организм быстро погибает, если не провести дефибрилляцию.
Ревербератор в однородной двухмерной системе возникает, например, при наличии отверстия, периметр которого превышает длину волны l = Vt (V – скорость) распространения волны.
Циркуляция волны возбуждения (ревербератор) возможна и в непрерывной среде, неоднородной по рефрактерности. Если в некоторую область среды, имеющую форму кольца, послать импульс, то он будет циркулировать, т.к. волны возбуждения, распро-страняющиеся по часовой стрелке и против нее, встретятся и погасят друг друга. Если рефрактерность не всюду одинакова, то циркуляция возникает вследствие трансформации (изменения) ритма.
27. МОДЕЛИРОВАНИЕ БИОФИЗИЧЕСКИХ ПРОЦЕССОВ
Метод моделирования является универсальным, применяемым практически во всех областях человеческой деятельности: науке, технике, экономике, медицине, архитектуре. Моделирование – основной метод био-физики. Он применяется на всех уровнях изучения живых систем, начиная от молекулярной биофизики, биофизики мембран, клетки, органов и кончая биофизикой сложных систем. Некоторые модели нами изучены в курсе биофизики. Это уравнение Ходжкина – Хаксли, модель мышечного сокра-щения, кровеносной системы и др.
При изучении сложных систем исследуемый объект может быть заменен другим – более простым, но сохраняющим основные, наиболее существенные для данного исследования, свойства. Такой объект исследования называется моделью. Модель – это всегда некоторое упрощение объекта исследования (его структуры, сложности связей, как внутренних, так и внешних).
На моделировании базируется любой метод научного исследования (теоретического, прикладного).
27.1. Этапы моделирования
1. Первичный сбор информации.
2. Постановка задачи и цели.
3. Обоснование основных допущений.
4. Создание модели, ее исследование.
5. Оценка адекватности модели реальному объекту. Границы применимости модели.
Модели бывают:
1. Физические (модель течения крови).
2. Биологические (гигантский аксон кальмара).
3. Математические (система дифференциальных уравнений).
27.2. Модель естественного роста численности популяции
Реальная система: Имеется некоторая популяция
Постановка задачи. Найти законы изменения численности популяции во времени.
Допущения:
1. Хищники отсутствуют.
2. Внутри вида нет борьбы за место обитания.
3. Размножение и естественная гибель, скорость их пропорциональна численности особей в данный момент.
Модель:
x – численность популяции в момент t.
R – скорость размножения.
g – коэффициент размножения.
S – скорость естественной гибели.
dx/dt – скорость изменения численности популяции.
e – коэффициент роста.
R = gx, S = – sx.
Уравнение баланса
Изменение численности особей в единицу времени определяется количеством рожденных за это время и умерших:
dx/dt =(g -s)х или dx/dt =eх
Начальное условие:
при t=0 x =xо
Интегрируем
х t
ò (dx/dt) =ò e dt
хо о
ln(x/xo) = et
x = xo e et
| e < 0 | e >0 |
 x
xo
t
x
xo
t
| x xo t |
Рис.54. Изменение численности популяции во времени
Анализ
1. e < 0 (при s >g) – скорость гибели больше скорости размножения, численность падает до нуля.
2. e > 0 (при g >s) – скорость размножения больше скорости гибели. Неограниченный рост со временем.
3. e = 0 (g =s). Численность неизменна.
Модель при e > 0 адекватна реальности лишь до определенного вре-мени. При большом количестве особей появляется борьба внутривидовая за место обитания – пищевые и водные ресурсы, генетический отбор.
27. 3. Модель изменения численности популяции
с учетом конкуренции между особями
Усложненная модель. Пусть существует внутривидовая борьба между особями, например, за место обитания.
Пусть скорость гибели за счет конкуренции пропорциональна вероятности встречи двух особей, тогда
S = –dx x – sx,
где s- коэффициент пропорциональности.
Уравнение баланса
dx/dt = gх – sх – dх2
тогда, dx/dt=eх – dх2 (нелинейное дифференциальное уравнение).
Заменяя U= (e – dх)/х, с учетом, что при t = 0 x = xo , получим
ln(x/xo) – ln[(e – dx)/(e – dxo)] = et,
отсюда
x(t) = {(xo/e)/[(e – dxo)e –et +dxo]}
при t ® ¥ х ® хст, хст = e/d,
где х ст – стационарный уровень.
 х
хст
хо
t х
хст
хо
t
| Модели 1 и 3 являются осно-вой моделирования процессов в биотехнологии (например, для установления оптимальных режи-мов выращивания различных микроорганизмов), развития эпи-демий. |
| Рис.55. Изменение численности популяции во времени с учетом конкуренции между особями |
27.3. Модель “ хищник – жертва”
Пусть живут два вида особей: зайцы (жертвы) и рыси (хищники).
Зайцы питаются растительной пищей (пищи достаточно), внутривидо-вая борьба отсутствует.
Рыси питаются только зайцами.
х – число жертв в момент времени t.
y – число хищников в момент времени t.
Уравнение баланса между численностью рожденных и гибнущих особей.
Жертвы:
dx/dt = gx - sx - axy
 |
gx - скорость размножения, sx - скорость естественной гибели, axy- скорость гибели за счет встречи с хищником.
Хищники:
dxy – скорость размножения, by – скорость естественной гибели.
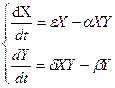 (27.1)
(27.1)
Находим стационарное решение x = const, y = const:
dx/dt =0 dy/dt =0.
Система дифференциальных уравнений сводится к алгебраическим:
хст(e - aуст) =0;
yст (dхст - b) =0.
Решения:
х ст =b/d;
Yст =e/a.
Предполагая, что произошли малые отклонения численности хищников V(t) и жертв U(t) относительно стационарных значений, имеем
х = хст +U(t), U<x ст , U <Yст.
Y =Yст +V(t), V <Yст, V < Xст.
dU/dt = Xст (e – aYст) + U (e – aYст) – aXст V – aUV,
dV/dt = Yст (d Xст – b) + V (dXст – b) + dU Yст +dUY
или
dU/dt = X 2ст [– aV/Xст – (aU/Xст)(V/Xст)],
dV/dt = Y2ст [d(U/Yст) + (d U/Yст)(V/ Yст )].
Пренебрегая членами второго порядка, с учетом (27.1) получим систему уравнений:
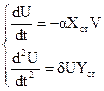 , (27.2)
, (27.2)
которую сводим к дифференциальным уравнениям второго порядка
d2U/dt2 +ebU = 0
и
d2V/dt2 +ebV = 0.
Это уравнения колебательных процессов.
Решения уравнений:
U = Umax sin 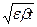 , V = Vmax sin( +j o)
, V = Vmax sin( +j o)
Амплитуды относятся:
Vmax /Umax = d/a Öe/b.
В результате численности особей при малых отклонениях от стационарных значений равны:
X(t) = Xст + Umax sin ;
Y(t) = Yст + Vmax sin( +j o )
Таким образом, численности популяций X и Y изменяются по синусо-идальному закону, колеблются около стационарных значений с частотой w = (e b)1/2 и смещены по фазе на jo. Обнаружена очень большая сходимость с опытными данными за период с 1845 по 1935 годы. Периодичность наблюдается примерно через 10 лет со смещением по фазе примерно на 1,5 года.
27.4. Фармакокинетическая модель
Для описания кинетики изменения концентрации введенного в организм лекарственного препарата предложена фармакокинетическая модель.
Необходимо найти законы изменения концентрации лекарственного препарата при различных способах и параметрах его введения и выведения.
В реальных условиях ввод и вывод лекарства сопровождаются большим числом разнообразных процессов. Это процессы всасывания в кровеносное русло при внесосудистом введении, перенос лекарства из крови к органам, удаление препарата из крови почками и другими органами.
Допущения:
1. Не рассматривается система органов, через которые последо-вательно проходит лекарство. Исключается многостадийность процессов ввода, переноса, вывода лекарственного вещества.
2. Не учитывается молекулярные механизмы процессов (проница-емость вещества, химические превращения).
3. Процессы ввода и вывода определяются скоростью.
Модель:
Существуют различные способы введения лекарственного вещества в организм. Законы изменения концентрации лекарственного вещества во времени C(t):
Однократное введение лекарственного препарата. Пациенту сделан укол.
 mo
Р
Организм mo
Р
Организм
| Представим себе орга-низм как систему объемом V. После введения в эту систему лекарственного пре-парата массой mo, начина-ется его удаление из орга-низма по схеме (рис.56). |
| Рис. 56. Однократное введение лекарствен- ного препарата |
Распределение препарата по объему предполагается равномерным. Скорость удаления препарата Р из организма прямо пропорциональна его массе в организме:
P = – km
k – коэффициент удаления препарата из организма.
Скорость изменения массы лекарственного вещества в организме (dm/dt) равна скорости его выведения Р:
(dm/dt) = Р, и, следовательно, (dm/dt) = – km.
Решение этого дифференциального уравнения с учетом начального условия при t = 0, масса введенного лекарства m = mo.
m = mo e -kt.
Концентрация лекарственного препарата в организме (например, в крови) c = m/V.
c = (mo/V) e –kt или С = Со e –kt,
где V – объем крови; Со – начальная концентрация.
С
 Со
t
Со
t
| Концентрация лекарственного препарата в крови будет непре-рывно снижаться по убывающему экспоненциальному закону (рис.57). При однократном способе введения лекарства не удается поддерживать в крови его постоянную концентрацию. |
| Рис.57. Изменение концентрации лекарства в организме при однократном способе введения |
Второй способ. Непрерывное введение препарата с постоянной скоростью. Пациенту поставлена капельница.
В этом случае изменение массы лекарственного препарата в организме dm/dt определяется не только скоростью его удаления Р, но и скоростью введения (рис.58). Q – количество лекарственного вещества, введенного в организм за единицу времени:

Q Организм Р
Рис.58.Непрерывное введение препарата с постоянной скоростью
(dm/dt) = Q – km.
Получим решение с учетом того, что при t = 0, m = 0:
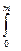 [ dm/(Q – km)] =
[ dm/(Q – km)] = 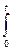 dt.
dt.
Введя новую переменную U= Q – km,
dU = – km, dm = – dU/k.
ò [dm/(Q – km)] = –(1/k)ò dU/U,
получим
ln (Q – km)/Q = –kt
и, наконец, –
m (Q/k)(1 – e –kt).
Концентрация лекарства в крови
C = (Q/kV)(1 – e –kt).
В начальный момент времени, при t = 0, С = 0, t ® ¥
e –kt ® 0 и С ® Q/kV.
Через некоторое время после начала введения лекарства устанавливается постоянная концентрация:
С = Q/kV.
Подбором скорости введения лекарств Q = kVСопт добиваются, что через некоторое время устанавливается оптимальная концентрация Сопт, которая необходима для терапевтического эффекта. При непрерывном способе введения лекарства удается достигнуть заданного результата С = Сопт только через некоторое время (рис.59).
 С
Сопт
t С
Сопт
t
| Оптимальная концентрация мо-жет быть установлена мгновенно при сочетании первого и второго спосо-бов. |
| Рис.59. Достижение оптимальной кон- центрации лекарственного препарата в организме при непрерывном введении |
Третий способ. Состояние непрерывного введения лекарственного препарата (первый способ) с введением нагрузочной дозы (второй способ).
 Сопт
t
Сопт
t
| m0 QP |
| Рис.60. Комбинированный способ введения лекарственного препарата |
В этом случае фармакокинетическая модель имеет вид
С=(Q/kV) – (1/V)(Q/k – mo)e –kt.
С = Сопт.
Если скорость введения лекарства (капельницей)
Q=kVCопт
и нагрузочная доза
mo=(Q/k)=VCопт,
то постоянная концентрация устанавливается мгновенно.
Фармакокинетическая модель дает возможность рассчитать оптимальное соотношение между параметрами ввода и вывода препарата для достижения терапевтического эффекта.
Дата публикования: 2015-11-01; Прочитано: 288 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
