 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Список источников
|
|
1. Наследственные болезни: национальное руководство/ Под ред.
акад. РАМН Н.П.Бочкова и др. – М.: ГЭОТАР-Медиа, 2012.- 936 с.
2. Геномика – медицине: Научное издание/ Под ред. академиков РАМН В.И.Иванова и Л.Л.Киселева. – М.: ИКЦ Академкнига, 2005. – 392 с.
3. Тарантул Б.З. Геном человека: Энциклопедия, написанная четырьмя буквами. Языки славянской литературы. — М., 2003. - 396 с.
4. Ormond K.E., Wheeler M.T., Hudgins L. et al. Challenges in the clinical application of whole-genome sequencing //Lancet. — 2010, May 15. - N 375(9727). - P. 1749-1751.
5. Samani N.J„ Tomaszewski М., Schunkert H. The personal genome — the future of personalised medicine? // Lancet. - 2010, May 1. - N 375(9725). - P. 1497-1498.
6. Wang E.T., Sandberg R., Luo S. et al. Alternative isoform regulation in human tissue transcriptomes // Nature. - 2008, Nov. 27. - N 456(7221). - P. 470-476.
7. Yoav Gilad, Victor Wiebe, Molly Przeworski, Doron Lancet, Svante Paabo. Loss of Olfactory Receptor Genes Coincides with the Acquisition of Full Trichromatic Vision in Primates // PLoS Biol. - 2004, Jan. - N 2(1). - P. 0120-0125.
Тема № 6. Этногеномика и геногеография
Краткое содержание:
1. Основные подходы к ДНК-анализу в популяционных исследованиях
2. Африканское происхождение человека современного типа
3. Использование анализа ДНК для изучения истории этносов
4. Этногеномика и геногеография Восточно-Европейского региона
5. Особенности русского генофонда
Последние годы на рубеже двух столетий ознаменованы стремительным прогрессом в области молекулярной генетики человека. Это связано, прежде всего, с работами по расшифровке генома человека, проведенными в рамках международных и национальных программ «Геном человека».
На основе этих исследований возникло новое научное направление, получившее название геномика, которое революционизировало всю современную биологию. В рамках самой геномики стали развиваться специализированные разделы: функциональная геномика, сравнительная геномика, медицинская геномика, компьютерная геномика и, наконец, наиболее захватывающий раздел — этническая геномика (этногеномика).
Основной задачей этногеномики является изучение геномного разнообразия в генофонде отдельных популяций, этносов, рас. Благодаря этнической геномике молекулярная генетика стала оказывать влияние не только на родственные разделы биологии и медицины, но и на такие отдаленные гуманитарные дисциплины как, например, история, антропология, включая происхождение, эволюцию, пути миграции, оценку родства и взаимодействия различных человеческих популяций.
1.Основные подходы к ДНК-анализу в популяционных исследованиях
Популяционные генетики обычно используют в работе различного рода полиморфные (вариабельные) признаки. Раньше в качестве таких признаков использовались полиморфные белки, например антигены групп крови, антигены главного комплекса гистосовместимости и др.
Сейчас эти маркеры стали называть классическими, чтобы отличить их от появившихся в последнее время маркеров ДНК. С помощью классических маркеров получено довольно много интересных сведений о популяциях различных регионов мира. Но подлинный переворот в этой области произошел при появлении нового инструмента в виде огромного числа полиморфных маркеров ДНК.
Преимущества изучения генетического разнообразия на уровне ДНК, по сравнению с белковым уровнем, колоссальны. Информационное содержание ДНК значительно выше белкового, особенно если сравнивать с той группой белков, которая доступна для массового анализа. Кроме того, техника исследования маркеров ДНК сводится к ограниченному числу методов, что позволяет автоматизировать процесс.
Важно отметить, что все маркеры ДНК с точки зрения популяционных исследований можно разделить на три группы: маркеры митохондриальной ДНК, маркеры Y-хромосомы и маркеры других хромосом.
В клетках человека, как и любого высшего организма, содержится геномы двух типов. Один находится в клеточном ядре и распределен по хромосомам — по 22 парам так называемых аутосом и по половым хромосомам — XX у женщин и XY у мужчин. В каждой паре хромосом (гомологичных) одна происходит от одного родителя, вторая — от другого.
Другой геном, совсем небольшой, принадлежит митохондриям (их считают энергетическими станциями клеток). Каждая митохондрия имеет свой геном, число митохондрий в клетке весьма велико. Вариабельность митохондриальной ДНК используется в популяционных исследованиях сравнительно давно из-за относительной простоты выделения ДНК. Основной особенностью этой вариабельности является материнский тип наследования.
Y-хромосомный полиморфизм является как бы зеркальным митохондриальному и имеет отцовский тип наследования. Оба типа вариабельности дополняют друг друга, давая раздельную информацию об отцовском и материнском вкладе в эволюцию популяций. Это явление дает новые, не существовавшие ранее, возможности в популяционных исследованиях — проследить и сопоставить историю женской и мужской части популяции и оценить их вклад в популяционный генофонд.
В отличие от них, ядерные маркеры ДНК, расположенные на других хромосомах, характеризуют сообщества людей в целом, не учитывая особенности генетического вклада различных полов.
Самой первой обнаруженной вариабельностью ДНК была вариабельность, связанная с точковыми заменами нуклеотидов (SNP), когда один нуклеотид в цепи ДНК меняется на другой.
Позднее был обнаружен другой тип полиморфизма, связанный с так называемыми гипервариабельными участками генома.
Эти участки содержат короткие последовательности ДНК, многократно повторенные один за другим стык в стык, так называемые тандемные повторы. Оказалось, что число повторений этих последовательностей сильно различаются у разных людей. У одного человека в конкретной точке генома последовательность может быть повторена 4 раза, а у другого 20, 40 и более раз. Из-за столь большого индивидуального разнообразия информативность этих маркеров очень высока.
Среди гипервариабельных участков, представленных тандемными повторами, выделяют мини - и микросателлиты. Для минисателлитов характерны более длинные элементарные звенья — от 10 и более нуклеотидов. Длина элементарного звена у микросателлитов меньше 10 нуклеотидных пар (н. п.). В основном исследователи имеют дело с микросателлитами с размером звена от 2 до 4-5 н.п. Полиморфизм мини- и микросателлитных локусов (участков хромосомы) довольно высок, отсюда следует, что и их информативность также должна быть весьма значительной.
Второй важной характеристикой маркеров ДНК является возможность оценки с их помощью временных параметров появления новых вариантов. Полиморфизм, отражающий замены единичных нуклеотидов, особенно в ядерных ДНК, основывается на событиях, происходящих достаточно редко, поэтому на основании полиморфизма данного типа можно судить о более отдаленных событиях популяционной истории.
Мини- и микросателлиты имеют более высокую скорость мутации и на основании этого они рекомендованы для анализа менее отдаленных этапов генетической истории.
2. Африканское происхождение человека современного типа
Первым полиморфизмом ДНК, широко использованным в популяционной генетике, явился полиморфизм митохондриальной ДНК. Дело в том, что в тот период еще не было метода полимеразной цепной реакции, а гены тестировали с помощью сложных и громоздких методов. Существенным было и то, что число копий митохондриальной ДНК (мтДНК) в клетке составляет от нескольких сотен до несколько тысяч. И, таким образом, этот материал мог быть более надежно тестирован, чем любая ядерная ДНК.
Необходимо напомнить коротко основные черты строения мтДНК. Это кольцевая двухцепочечная молекула, у человека ее размер составляет 16569 пар оснований. Основная часть полиморфизма мтДНК связана с небольшим районом в 1,2 kb, называемым контрольным районом. Здесь содержатся последовательности, контролирующие транскрипцию и репликацию. Этот район известен также как D-петля (displacement — перестройка). Он высокополиморфен и содержит два гипервариабельных региона, примерно по 400 bp. В обоих регионах содержится большое количество точковых замен.
Таким образом, эти участки анализируют в виде гаплотипов (сочетаний вариабельных участков), число вариантов которых в популяциях очень велико.
Напомним, что митохондрии наследуются по материнской линии, так как в оплодотворенное яйцо они попадают из яйцеклетки. Судьба небольшого числа единичных митохондрий сперматозоида, которые могут оказаться в оплодотворенной яйцеклетке, неизвестна — во всяком случае, они не проявляют себя в новом организме. Таким образом, анализ мтДНК дает информацию о генетической истории по женской линии человечества.
Изучение вариантов митохондриальных ДНК в различных популяциях мира показало, что все они могут быть выведены из одного единственного варианта. Эта работа, выполненная в конце 1990-х гг., вызвала большой резонанс, в ней сформулировано представление о митохондриальной Еве, прародительнице всего человечества.
Одновременно изучался и генетический материал Y-хромосомы, в том числе в строение полиморфных маркеров. В нерекомбинирующей области Y-хромосомы обнаружено множество полиморфных маркеров, образующих гаплотипы, т. е. сочетания вариабельных участков. Такие гаплотипы в нерекомбинирующей области Y-хромосомы, обладающие большой устойчивостью во времени, используют в качестве инструментов для изучения давних генетических событий, в особенности миграций.
То, что многообразие ДНК африканских популяций выше, чем всех остальных, было показано не только с помощью митохондриальных маркеров, но и впоследствии с помощью ядерных, включая Y - хромосомные.
Наиболее древние варианты Y - хромосомы найдены в ряде популяций Африки, в частности у койсанов. Таким образом получается, что и Адам — прародитель нашего рода — выходец из Африки.
Большая работа была проведена также и по маркерам других ядерных хромосом. Все эти данные подтвердили африканское происхождение всего человечества.
Многочисленные исследования показали, что все человеческие митохондриальные ДНК могут иметь единого предка, и с некоторыми допущениями можно рассчитать, когда произошло первое ветвление генеалогического древа митохондриальной ДНК. Важным условием для этого является знание скорости мутаций.
Одним из подходов к калибровке митохондриальных часов является сравнение этих последовательностей для человека и шимпанзе, с учетом того, что эти виды разошлись друг от друга 5-7 миллионов лет назад. Оценка средней скорости мутаций в ДНК митохондрий была проведена в нескольких исследованиях и составила (1-5)х10-6 мутаций на нуклеотид на поколение, что по крайней мере на два порядка выше, чем скорость мутаций в ядерной ДНК.
Расчеты, проведенные на основе этих результатов, показали, что расхождение митохондриальных ДНК началось около 150 тысяч лет назад. Первое «расхождение» вариантов митохондриальных ДНК произошло в древности внутри африканского континента, дав начало трем родословным. Расселение по другим континентам осуществлялось потомками только одной из трех африканских ветвей. Самая древняя миграция проходила по южному побережью Азии, через Новую Гвинею — в Австралию примерно 70 тыс. лет назад. Следует отметить, что в это время Австралия, Тасмания и Новая Гвинея были в составе единого материка.
Интересно, что из-за сниженного уровня моря в то время Малайский полуостров, острова Суматра, Ява, Борнео и Бали также были едины. Все это значительно облегчало продвижение людей с южного побережья Азии в Австралию. Европа, согласно этим данным, заселялась позднее, что, по-видимому, было связано с более суровыми климатическими условиями и наличием здесь неандертальцев, хорошо приспособленных к холодному климату.
В этой связи интересное исследование было проведено на мтДНК, выделенных из костей неандертальцев. Один из образцов — это известная находка из Дюссельдорфа, обнаруженная в 1856 г. Было расшифровано 380 нуклеотидов из первого гипервариабельного региона (ГВС1) D-петли. Если средние попарные различия у современного человека в этой области составляют величину 8,0 (с колебаниями от 1 до 24), то размах различий между неандертальцем и современным человеком составил от 22 до 36. Общий предок для этих подвидов, как показали расчеты, может быть датирован в пределах от 550 до 680 тысячелетий тому назад.
Было проведено исследование ДНК других неандертальских образцов в сравнении с древним образцом ДНК кроманьонца. Полученные данные продемонстрировали геномные различия неандертальца и кроманьонца и явились дополнительным подтверждением того, что это, очевидно, разные подвиды одного вида Homo sapiens.
Исследование Y-хромосомной вариабельности в глобальном масштабе было недавно проведено Петером Андерхиллом, одним из сотрудников Кавалли-Сфорца. Был проведен анализ 166 полиморфных точек в Y-хромосоме более 1000 мужчин из разных регионов Земли. В результате обнаружено 116 гаплотипов, представляющих собой отдельные исторические родословные, которые удалось объединить в одно эволюционное древо. В этом древе выделились 10 ветвей, каждая из которых соответствует конкретным географическим регионам.
В Африке обнаружены варианты Y-хромосомы, соответствующие трем ветвям, причем первая из них является самой древней и несет в себе некоторые особенности, общие с нашими ближайшими «родственниками» — приматами. Эта ветвь встречается у некоторых африканских меньшинств — у койсанов, у ряда популяций суданцев и эфиопов. Все остальные ветви отличаются от ветви N1, и они, собственно, и образуют основной «ствол» данного древа. Вторая и третья ветвь являются тоже африканскими, причем третья ветвь особенно широко представлена у разных народов континента. Именно эта ветвь наиболее родственна Y-хромосомным вариантам всего остального человечества. Интересно, что одной из ветвей, наиболее близкой к африканским, является австрало-новогвинейская, а наиболее отдаленной — ветвь американских индейцев. Если сравнить эти результаты с данными по митохондриальной ДНК, то можно видеть, как они хорошо согласуются друг с другом. Такое согласование говорит о том, что полученные сведения отражают реальный эволюционный путь современного человека, независимо записанный в родословных как по женской, так и по мужской линии.
Далее были проведены исследования по различным типам полиморфизма ядерной ДНК других хромосом. Оказалось, что все они пригодны для оценки путей миграции и даже (в первом приближении) — времени, когда данное событие происходило. Особенно подходящими для этих целей оказались гаплотипы, состоящие из сочетаний близко расположенных маркеров различного типа. Они явились особенно полезными при анализе происхождения популяций и реконструкции исторических миграционных процессов.
Для множества генов были исследованы гаплотипы, составленные из полиморфных участков. Были изучены десятки популяций из различных географических регионов. Оказалось, что наибольшее разнообразие гаплотипов имеется в африканских популяциях, проживающих южнее Сахары. Все остальные изученные популяции мира выглядели как одна из подгрупп африканцев.
Эти данные показали, что популяции Северо-Востока Африки в ранней истории отделились от остальных африканских популяций, после чего часть из них мигрировала из Африки на другие континенты. Многие показатели, выявленные в данных работах, позволяют считать, что африканские популяции име больший эффективный размер и высокий уровень полиморфизма.
Таким образом, исследование геномного разнообразия человека убедительно показало, что все человечество имеет единое происхождение и ведет свой род из Африки. Все три независимые линии анализа — с помощью митохондриальной ДНК, маркеров Y-хромосомы и ядерных маркеров других хромосом привели к одним и тем же результатам, доказавшим наше африканское происхождение.
3. Использование анализа ДНК для изучения истории этносов
Молекулярно-генетические подходы эффективны не только при изучении глобальных вопросов эволюции человека как вида. Большую роль маркеры ДНК играют и при изучении этнической истории в отдельных регионах мира. Один из весьма изученных регионов — это Западная Европа.
В работе Джауме Бертранпетита и его коллег был проведен анализ митохондриальной ДНК из популяций Европы и Ближнего Востока. Всего было исследовано около 500 человек, среди них — баски, британцы, швейцарцы, тосканцы, сардинцы, болгары, турки, жители Ближнего Востока, включавшие бедуинов, палестинцев и йеменских евреев — т. е. народов, относящихся к европеоидам. В данной работе, как и во многих предыдущих, был продемонстрирован низкий уровень генетического разнообразия европейцев, по сравнению с другими, в особенности, африканцами. Это может быть связано с разными причинами: например, с относительно недавним их происхождением, с высокой скоростью миграции, или в связи с быстрым демографическим ростом, который, как полагают, происходил в доледниковый период.
Однако, несмотря на сравнительную гомогенность европейских популяций, имеются определенные географические различия в распределении наблюдаемой генетической вариабельности. Это позволило достоверно реконструировать пути миграции народов в далеком прошлом.
Полученные результаты подтвердили предположение о передвижении населения из Ближнего Востока в Европу. Расчеты показали, что эта миграция осуществлялась в течение длительного времени — на протяжении десятков тысячелетий. Данные позволяют предположить, что основные генетические характеристики европейцев, по-видимому, сложились еще в палеолите, тогда как более поздние неолитические миграции оказывали меньшее влияние на изучаемый генофонд.
К аналогичному выводу пришли и другие исследователи, проведя анализ митохондриальных ДНК у более чем 700 человек из 14 популяций Европы и Ближнего Востока. Подробный анализ ветвей каждого варианта мтДНК позволил авторам сделать следующий вывод: большинство населения современной Западной Европы, является потомками ранних поселенцев, пришедших из районов Ближнего Востока в период верхнего палеолита. Обнаружены также «следы» и более поздних продвижений в Европу выходцев из Ближнего Востока, однако эти миграции оказали значительно меньшее влияние, чем предыдущая.
В последующих работах, выполненных Торони и коллегами, были также исследованы митохондриальные ДНК жителей Европы, Ближнего Востока и северо-западной Африки. При этом в каждом образце был осуществлен анализ обоих гипервариабельных участков, а также полиморфизма вдоль всей молекулы, что позволило определить гаплотип в каждом образце и выявить родственные группы гаплотипов, обозначенные как
гаплогруппы.
Эти исследования показали, что у европейцев с наибольшей частотой встречаются две родственные гаплогруппы митохондриальной ДНК, обозначенные авторами как Н и V. Подробный анализ этих гаплогрупп, включая их географическое распределение, позволил авторам сделать предположение, что гаплогруппа V является автохтонной (т. е. местной) для Европы. Она возникла 10-15 тысячелетий назад на севере Иберийского полуострова или на юго-западе Франции, затем диффундировала на северо-восток (вплоть до Скандинавии) и на юг до северо-запада Африки.
В настоящее время она с наибольшей частотой встречается у басков и саамов (которые считаются самыми древними жителями Европы), но отсутствует на Кавказе, юге Европы и Ближнем Востоке. Оценка среднего числа нуклеотидных различий от предкового гаплотипа показывает, что иберийские популяции имеют наибольшее разнообразие по данному признаку. Именно это позволило сделать вывод, что с большой вероятностью местом возникновения группы V является Иберийский полуостров и примыкающие к нему территории юго-западной Франции.
Гаплогруппа Н является самой распространенной в Европе, она встречается в разных популяциях с частотой от 20 до 60%, обнаруживая постепенную (клинальную) изменчивость с востока на запад и север. Она обнаруживается с меньшей частотой в других европеоидных популяциях, например, на Ближнем Востоке, в Индии, на севере Африки, в Сибири. Интересно, что наибольшее разнообразие вариантов гаплогруппы Н обнаружено в популяциях Ближнего Востока. Это позволяет считать, что она возникла именно в этих популяциях, причем оценка ее возраста составляет 25-30 тысячелетий. Однако в Европу она проникла позднее — 15-20 тысячелетий назад, т. е. в период верхнего палеолита.
Таким образом, данная работа выявила множество интересных деталей в генетической истории европейцев, но в целом подтвердила прежние результаты о древности этих популяций (по крайней мере, по женской линии).
Изучение полиморфизма Y-хромосомных маркеров у европейцев также показывает их древнее происхождение. Работа Семино и соавторов так и называется: «Генетическое наследие человека палеолита в ныне живущих европейцах: возможности Y-хромосомных маркеров». В этой работе принимал участие большой интернациональный коллектив, состоящий из двух американских и нескольких европейских лабораторий, включая российскую. Было изучено более 1000 мужчин из 25 разных регионов Европы и ближнего Востока.
Анализ по 22 маркерам Y-хромосомы показал, что более 95% изученных образцов могут быть сведены к десяти гаплотипам, т. е. к 10 историческим родословным. Из них два гаплотипа, обозначенные как Eu 18 и Eu 19, появились в Европе в палеолите. Более 50% всех изученных европейских мужчин относятся к этим древним гаплотипам. Они являются родственными и отличаются лишь одной точковой заменой (мутация М17), однако их географическое распределение имеет противоположную направленность. Частота Eu18 уменьшается с запада на восток, будучи наиболее выраженной у басков. Оценка возраста этого гаплотипа составляет примерно 30 тысяч лет, возможно, это самая древняя родословная в Европе. По типу географического распределения она очень напоминает распределение митохондриальной гаплогруппы V, также имеющей верхне-палеолитическое происхождение. Можно предположить, что гаплотип Eu18 Y-хромосомы и гаплотип V митохондриальной ДНК являются характеристиками одной и той же древней европейской популяции, проживавшей в верхнем палеолите в районе Пиренейского полуострова.
Родственный Y-хромосомный гаплотип Eu19 имеет совсем другое распределение в европейских популяциях. Он отсутствует в Западной Европе, его частота увеличивается к востоку и достигает максимума в Польше, Венгрии и на Украине, где предыдущий гаплотип Eu18 практически отсутствует. Самое высокое разнообразие микросателлитных маркеров в составе гаплотипа Eu19 найдено на Украине. Это позволило сделать предположение, что именно отсюда началась экспансия данной исторической родословной. К сожалению, среди вариантов митохондриальной ДНК пока не найдено такого, который имел бы сходное с Eu19 географическое распределение.
Как можно объяснить столь различную картину распространения столь родственных гаплотипов? Из данных по распространению Eu18 и Eu19 можно предположить, что это связано со следующим сценарием. Во время последнего ледникового периода люди вынуждены были покинуть Восточную и Центральную Европу. Часть из них переместилась в Западные области. Некоторые нашли убежище на Северных Балканах, единственном месте в Центральной Европе, где была возможность существования. Таким образом, ледниковый период люди переживали в 2-х регионах (западная Европа и Северные Балканы), находясь в значительной изоляции друг от друга. Такой сценарий подтверждают также данные по флоре и фауне того же периода. Здесь также была выявлена изоляция в указанных областях в ледниковый период. После чего наблюдалось распространение переживших видов и популяций из данных заповедных мест.
Дополнительные молекулярно-генетические данные подтверждают наличие двух очагов, из которых происходило распространение двух рассмотренных гаплотипов.
Среди других Y-хромосомных гаплотипов большая часть имеет географическое распределение, указывающее на их происхождение из региона Ближнего Востока. Однако два из них появились в Европе (или, возможно, здесь и возникли) в палеолите.
Характеристики этих исторических родословных очень напоминают таковые для гаплогруппы Н митохондриальной ДНК. Возможно, что они маркируют одни и те же исторические события, связанные с расселением ближневосточных популяций в Европе в период, предшествующий последнему ледниковому максимуму.
Все остальные Y-хромосомные гаплотипы появились в Европе позже. В неолите произошло распространение ряда гаплотипов из региона Ближнего Востока, по мнению многих авторов, в связи с распространением земледельческой культуры.
Интересно, что в работе был выявлен новый вариант Y-хромосомы (мутация М178), встречающийся только в северо-восточных областях Европы. Возраст этого гаплотипа оценивается величиной, не превышающей 4000 лет, а его распространение может отражать сравнительно недавнюю миграцию уральских популяций.
Таким образом, в данной работе показано, что лишь немногим более 20% европейских мужчин относятся к историческим родословным (выявленным с помощью Y-хромосомного полиморфизма), которые появились в Европе сравнительно недавно — после ледникового периода в неолите. Около 80% мужчин Европы относятся к более древним европейским родословным линиям, нисходящим ко времени верхнего палеолита.
В последнее время активно дискутировалась идея, высказанная Марком Стоннекингом еще в 1998 году, что более высокая вариабельность популяций (особенно европейских) по Х-хромосомным маркерам, в сравнении с митохондриальными, связана с различиями в дистанциях миграций между женщинами и мужчинами. Согласно этой идее, миграция мужчин оказывается более ограниченной пространственно, чем миграция женщин. Однако к таким выводам следует относиться с большой осторожностью, так как еще многие популяционные свойства маркеров ДНК, особенно в сравнении одного с другим, мало изучены. Кроме того, большой вклад могут вносить в это явление социально-демографические факторы, например, такие, как полигамия, имеющаяся или имевшаяся ранее у многих народов.
Тем не менее, необходимо подчеркнуть, что наличие такой возможности, как анализ отдельно и мужской, и женской популяционной истории, открывает новые перспективы в изучении популяций, которых не было ранее, до обнаружения полоспецифических маркеров ДНК, связанных с митохондриальным и Х-хромосомным полиморфизмом.
Изучение популяций американских индейцев и их связи с сибирскими народами также осуществлялось с помощью маркеров ДНК. Проблема раннего заселения Американского континента представляет собой одну из наиболее противоречивых тем в исследованиях по эволюции человека. На основании данных антропологии, археологии, лингвистики и генетики принято считать, что предки коренного населения Америки прибыли из Азии. Однако время, место происхождения и число волн миграции до сих пор являются предметом дискуссий.
Ранее, на основании синтеза мультидисциплинарных исследований было высказано предположение о трех независимых волнах миграции предковых азиатских популяций через Берингов пролив. Изучение классических маркеров ДНК выявило тенденции, которые можно расценивать как подтверждение трехволновой модели миграции.
Однако первые результаты анализа митохондриальной ДНК показали, что их интерпретация может быть значительно шире, в том числе — в поддержку модели с четырьмя волнами миграции. Дальнейший анализ данных по митохондриальной ДНК позволил свести их к одному предположению, что все популяции американских индейцев могут быть сведены к единой предковой популяции, проживавшей ранее в регионе Монголии и Северного Китая.
Для того чтобы проверить столь противоречивые гипотезы, необходимо было исследовать дополнительные полиморфные системы ДНК. Было проведено исследование 30 вариабельных Y-хромосомных локусов у американских индейцев и нескольких сибирских популяций в сравнении с другими регионами мира. Это позволило выявить общих предков коренных жителей Америки с популяциями кетов из бассейна реки Енисей и с популяциями алтайцев, населяющих Алтайские горы. Таким образом, было показано преимущественно центрально-сибирское происхождение американских индейцев по мужской линии, которые могли мигрировать в Америку в доледниковый период.
Карафет и соавторы исследовали более 2000 мужчин из 60 популяций мира, включая 19 групп американских индейцев и 15 групп аборигенных сибирских народов. В данном исследовании было показано, что у американских индейцев имеется не один праотцовский гаплотип, а девять, причем два из них являются исходными, родоначальными гаплотипами Нового Света. Т.е. можно было предполагать по меньшей мере две волны миграции в Новый Свет, причем обе из региона озера Байкал, включая Саянские и Алтайские горы. И, наконец, самые последние данные однозначно показали, что была одна волна миграции из Сибири в Америку 13 тысяч лет назад.
С помощью полиморфных маркеров ДНК были проведены интересные исследования по заселению тихоокеанских архипелагов, а также острова Мадагаскар. Существовала точка зрения о переселении людей из Юго- Восточной Азии на тихоокеанские острова. Однако подробный анализ показал, что это был непростой и длительный процесс.
Изучение митохондриальных ДНК в данном регионе показало, что на островах Океании часто встречается (с частотой до 80-90%) специфическая делеция в 9 пар нуклеотидов, в Юго-Восточной Азии она встречается значительно реже. Подробный анализ показал, что данная делеция встречается в разном генетическом контексте, т. е. в сочетании с различными полиморфными участками. Эти сочетания принято называть мотивами, причем различают меланезийский, полинезийский и мотив Юго-Восточной Азии. Все представленные данные позволили предположить, что население островов Меланезии и Юго-Восточной Азии (Индонезия) в древности не смешивалось. Восточная Полинезия заселялась из обоих этих регионов очень малыми группами, что привело к формированию смешанного генофонда этих островов.
Интересной работой является исследование населения Мадагаскара, проводимое в течение многих лет Химлой Содиал и коллегами. История и время заселения этого острова остаются неизвестными из-за отсутствия письменных свидетельств. Немногочисленные археологические данные указывают, что первые поселенцы явились выходцами предположительно из Индонезии (находки датируются началом первого тысячелетия нашей эры), позднее датируется волна заселения из Африки. От Африки Мадагаскар отделен проливом шириной 400 км, расстояние до Индонезии — 6400 км. Население острова составляет 11 млн человек и подразделено на 18 этнических групп. В диалектах имеются особенности, указывающие на арабское и африканское влияние.
Изучение митохондриальной ДНК у населения Мадагаскара обнаружило высокую частоту специфической делеции размером 9 пар нуклеотидов, находящейся в окружении полиморфных участков, называемых полинезийским мотивом. Этот результат можно объяснить тем, что первые поселенцы Мадагаскара, по-видимому, были мореплавателями и прибыли из Полинезии или относились к той популяции, выходцы из которой заселяли Полинезию, но их путь в Мадагаскар проходил через Индонезию. То, что эти данные получены при анализе митохондриальной ДНК, говорит о том, что в составе прибывших на Мадагаскар групп имелись женщины.
Изучение Y-хромосомного полиморфизма у мужчин Мадагаскара показало следующую картину. Большая часть (более чем 2/3) современных родословных линий относится к африканскому типу и только 15% — к вариантам из Юго-Восточной Азии. Это говорит о том, что переселение из Африки, которое могло происходить как одновременно, так и в более позднее время, чем азиатское, осуществлялось бо́льшим числом людей. Было показано, что обе линии переселенцев, как африканских, так и азиатских, пережили период резкого снижения численности, возможно из-за каких-то внешних воздействий (природные аномалии, эпидемии чумы или что-то еще).
Очень интересное исследование, которое осуществляется несколькими интернациональными группами, ведется в Индии. Известна высокая подразделенность индийского общества, в том числе кастовая. Изучение митохондриальной ДНК и Y-хромосомного полиморфизма у представителей различных каст и племен выявило много любопытных деталей. Женское население Индии, как показывает данное исследование, выглядит более или менее гомогенным. Более 60% жителей Индии имеют варианты митохондриальной ДНК, относящиеся к древней группе ранней (возможно, первой) волне миграции из Восточной Африки, осуществлявшейся примерно 60 тыс. лет назад. В то же время в некоторых районах Индии ввысших кастах содержание вариантов митохондриальной ДНК, сходных с европейскими, выше, по сравнению с низшими кастами.
Что касается Y-хромосомного анализа, то здесь выявлены более четкие корреляции с кастовой принадлежностью. Чем выше ранг касты, тем выше содержание вариантов, сходных с европейскими, причем, что особенно интересно, с восточноевропейскими. Это является подтверждением точки зрения некоторых археологов, что прародина завоевателей Индии — индо-ариев, основавших высшие касты, находится на юге Восточной Европы.
Удивительные результаты были получены совсем недавно интернациональной группой под руководством английского исследователя Криса Тайлер-Смита. Проводилось широкомасштабное изучение Y-хромосомного полиморфизма во множестве азиатских популяций: в Японии, Корее, Монголии, Китае, в государствах Средней Азии, в Пакистане, Афганистане и на Южном Кавказе. В 16 популяциях из довольно обширного азиатского региона, простирающегося от Тихого океана до Каспийского моря, достаточно часто встречалась одна и та же генетическая линия Y-хромосомы. В среднем по данному региону эта линия встречается у 8% мужчин. Это составляет 0,5% всего мужского населения Земли. В некоторых районах внутренней Монголии, Центральной и Средней Азии данная линия встречается с частотой от 15 до 30%.
Расчеты показывают, что эта линия Y-хромосомы произошла в Монголии примерно 1000 лет назад (в интервале 700-1300 лет) и быстро распространилась по указанной территории. Такое явление не могло произойти случайно. Если бы причиной была миграция некой популяции, то исследователи должны были обнаружить несколько таких линий. Проанализировав географию распространения и время возникновения данной генетической линии, авторы сделали сенсационное предположение, что этот генетический вариант принадлежит Чингисхану и его ближайшим родственникам по мужской линии. В пределах обозначенного времени на данной территории действительно существовала империя именно этого завоевателя. Известно, что сам Чингисхан и его ближайшие родственники имели много потомков, которые сохраняли свое престижное положение на протяжении длительного времени. Таким образом, здесь происходил отбор не вследствие биологического преимущества, а по социальным причинам, что представляет собой новое явление в генетике.
Из приведенных примеров изучения популяций различных регионов мира видно, что маркеры ДНК дают новое понимание многих аспектов эволюции человека, как недавних, так и отдаленных.
4. Этногеномика и геногеография Восточно-Европейского региона
Народы Восточной Европы до недавнего времени оставались белым пятном в исследованиях по ДНК-полиморфизму. Имелись только отдельные разрозненные данные по некоторым популяциям. К настоящему времени российскими учеными проведено изучение восточноевропейских популяций по нескольким полиморфным локусам ДНК. Полученные результаты вызвали большой интерес.
Изученные полиморфные локусы представлены в основном мини- и микросателлитами, которые обладают расоводиагностическими свойствами, особенно в отношении европеоидных и монголоидных групп. Другими словами, распределение аллельных вариантов каждого маркера существенно различается у европеоидных и монголоидных народов.
В качестве примера можно привести вариабельный участок генома, имеющий особый интерес. Это ген рецепторного белка CCR5. Есть данные, что вирус СПИД использует именно этот рецептор для внедрения в клетки. Оказалось, что у некоторых людей в этом гене имеется нехватка (делеция) 32-х нуклеотидов. В результате свойства рецептора меняются таким образом, что он становится устойчивым к вирусу СПИД. При этом люди, получившие такую делецию от обоих родителей, не могут заразиться СПИД, даже находясь в группах высокого риска.
В Азии и в Африке эта делеция практически не встречается, но обнаруживается с разной степенью частоты в европейских популяциях, причем имеется три очага с максимумом и все они расположены в северной и восточной части Европы.
Совершенно непонятно происхождение этих очагов — ведь европейское население с вирусом СПИД ранее не встречалось. По-видимому, какие-то еще инфекционные агенты (возможно, вирус натуральной оспы) пользуются тем же рецептором для проникновения в клетки, и именно в связи с этими инфекциями произошла селекция, приведшая к накоплению данной делеции в обнаруженных очагах. Тем не менее, наличие или отсутствие делеции в данном гене также является одним из типов полиморфизма, который используется для анализа популяций. Другие маркеры, вариации в которых связаны с тандемными повторами, также обладали свойствами расоводиагностических признаков.
Суммированные результаты по всем локусам были проанализированы с помощью наиболее современного подхода компьютерной геногеографии. Разработка этого подхода была начата под руководством проф. Рычкова Ю. Г. и продолжена его учениками в Медико-генетическом научном центре РАМН и в Институте общей генетики РАН.
Были проведены исследования обобщающих характеристик генофонда восточноевропейского региона. Для этого использовали метод главных компонент в его картографическом варианте. Этот метод позволяет провести «сжатие» объемов статистической информации без потерь ее информативности. Это дает возможность объединить данные по всем изученным локусам и выявить основные закономерности изменчивости генофонда. Первая главная компонента включает в себя основные и наиболее общие закономерности в изменчивости изученных генов в пространстве региона. Вторая главная компонента и так далее — являются следующими по рангу и значимости величинами.
Геногеографические карты главных компонент наглядно отражают изменчивость всей совокупности генов и воспроизводят наиболее общие тенденции в географии генофонда в целом.
Родная Европа и чуждая Азия. Бросается в глаза (рис.1) близость практически всей Европы к нашей «среднерусской популяции». Широкая область светлых тонов фиксирует популяции, наиболее близкие к «среднерусским». Оказывается, что практически все популяции центральной Европы чрезвычайно близки к «среднерусским» значениям: величины генетических расстояний варьируют от 0 до 0.01. Во всей Центральной и Западной Европе от «среднерусских» значений резко отличается только северная часть Скандинавии, одна румынская и одна из множества итальянских популяций. Умеренные отличия показывают Франция, Англия, Испания и Италия - но, например, Германия, Австрия, Польша на карте неотличимы от среднерусской популяции. Однако даже генетические отличия испанцев заметны лишь в общеевропейском масштабе и исчезают, когда мы охватываем взглядом всю Евразию - повторимся, что на фоне Евразии вся Европа удивительно похожа на средний русский генофонд.
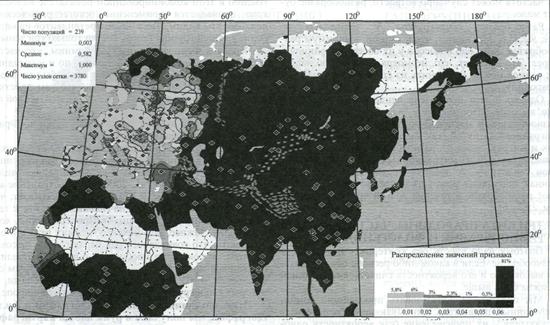
Рисунок 1- Карта генетических расстояний по мтДНК: от русского генофонда до популяций Евразии[2]
Главное, что обнаруживает карта генетических расстояний - это удивительную близость «среднерусских» значений к вариациям европейского
генофонда. Это сходство - весьма нетривиальный результат, особенно на фоне радикальных отличий генофонда всего остального евразийского населения. Резкий градиент проходит в районе Урала, пересекает Кавказ и далее на юге немного размывается, показывая «слегка русские» Анатолию и Левант. Все популяции к югу и востоку от этой линии обладают иным митохондриальным генофондом, непохожим на средний русский. Территории, расположенные западнее этой линии, за рядом исключений, достаточно близки к «усредненной русской популяции».
Европейцы больше русские, чем сами русские! Однако самое удивительное, что одним из таких исключений является часть русских популяций, белорусы и украинцы Поднепровья: они значительнее отличаются от «среднерусской популяции», чем многие иные популяции Европы. Это - неожиданный результат. Он показывает, сколь велик размах генетических различий в пределах русского генофонда, насколько значительно некоторые русские популяции могут отличаться от усреднённых общерусских величин. Одна зона слегка повышенных отличий от среднерусского генофонда расположилась прямо... в Центральной России! Другая - на северо-западе, хотя эти данные не слишком надёжны, так как основываются всего на одной псковской популяции. Третья - северная зона - особенно обращает на себя внимание. Она отмечает то - пока ещё умеренное - отличие северно-русского генофонда от генофонда усредненно-русского, которое далее переходит уже в резкие отличия финских, саамских и зауральских популяций.
Итак, русский этнос имеет сложную внутреннюю структуру. Карта генетических расстояний по частотам гаплогрупп мтДНК показывает порой большее сходство европейских групп с усреднённой русской популяцией, чем сами русские популяции: между русскими группами выявлены значительные различия.
Коренное население Азиатских регионов Евразии) резко отличается от русских популяций.
Дата публикования: 2015-10-09; Прочитано: 560 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
