 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Механизм мышечного сокращения
|
|
В соответствии с теорией скользящих нитей мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах, при этом тонкие актиновые нити скользят вдоль толстых миозиновых, двигаясь между ними к середине саркомера.
Миозиновые нити несут поперечные выступы с головками, состоящими примерно из 150 молекул миозина (рисунок 7.6А). Во время сокращения каждая головка (поперечный мостик) может связывать миозиновую нить с соседними актиновыми. Движение головок создаёт объединённое усилие, как бы «гребок», продвигающий актиновые нити в середину саркомера [31].

Рисунок 7.6 Функция поперечных мостиков.
А – модель механизма сокращения: миозиновая нить с поперечными мостиками, прикреплёнными к соседним актиновым нитям. Вверху – до, внизу – после «гребкового» движения.
Б – модель механизма генерирования силы при статическом напряжении. Слева - до, справа – после «гребка» (по Й. Рюэгг[31]).
В расслабленном состоянии механизм взаимодействия миозиновых головок с актиновыми нитями блокируется за счёт того, что участок актиновых нитей, предназначенный для связывания с головками миозина, перекрыт молекулами белков тропонина и тропомиозина.
Мышечное сокращение происходит под воздействием двигательного нервного импульса, представляющего собой волну повышенной мембранной проницаемости, распространяющейся по нервному волокну. Эта волна передаётся на Т-систему саркоплазматической сети и в конечном итоге достигает цистерн, содержащих ионы кальция. Проницаемость стенок цистерн повышается, ионы кальция выходят из цистерн в саркоплазму, где их концентрация за очень короткое время (около 3милисекунд) увеличивается примерно в 1000 раз. Ионы кальция, взаимодействуя с тропонином, изменяют его пространственную форму, что вызывает смещение молекул тропомиозина, обеспечивая доступ миозиновых головок к активным участкам актиновых нитей. В результате этого между миозином и актином возникает поперечный мостик, расположенный под углом 90о (рисунок 7.6А). Поскольку в толстые и тонкие нити входит большое число молекул миозина и актина, между мышечными нитями образуется довольно большое число поперечных мостиков. Образование связи между актином и миозином сопровождается повышением АТФ-азной активности миозина, разрешая её расщепление. За счёт энергии, выделяющейся при гидролизе АТФ, миозиновая головка подобно веслу лодки поворачивается и мостик между толстыми и тонкими нитями оказывается под углом 45о, что приводит к скольжению мышечных нитей навстречу друг другу (рисунок 7.6А) [11].
Совершив поворот, мостики между тонкими и толстыми нитями разрываются, АТФ-азная активность миозина резко снижается и гидролиз АТФ прекращается. Но если двигательный нервный импульс продолжает поступать в мышцу и в саркоплазме сохраняется высокая концентрация ионов кальция, поперечные мостики образуются снова, повторно включается гидролиз АТФ, дающий энергию для поворота поперечных мостиков с последующим их разрывом. За счёт ритмичных прикреплений и отделений миозиновых головок актиновая нить подтягивается к середине саркомера подобно тому, как группа людей тянет верёвку, перебирая её руками. Когда принцип «вытягивания верёвки» реализуется во множестве последовательно расположенных саркомеров, повторяющиеся молекулярные движения поперечных мостиков приводят к движению всей мышцы [31].
Каждый цикл сокращения, включающий образование, поворот и разрыв мостика, требует расходования одной молекулы АТФ в качестве источника энергии [11].
При сокращении мышцы происходит её укорочение. Но при статической нагрузке длина мышцы не изменяется. Что же в таком случае происходит в мышце с точки зрения «гребковой» теории Хаксли?
Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга. Процесс генерирования изометрической (т.е. статической) силы показан на рисунке 7.6Б. Сначала головка миозиновой молекулы прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом 45о, возможно благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика в напряжённое состояние. Упругое напряжение, создаваемое индивидуальным поперечным мостиком очень мало и для создания необходимой мышечной силы требуется объединение усилий огромного количества таких соединённых параллельно мостиков. Они будут тянуть соседние актиновые нити, как команда игроков тянет канат. Но даже при изометрическом сокращении поперечные мостики не находятся в непрерывно напряжённом состоянии. Каждая миозиновая головка уже через десятые или даже сотые доли секунды отделяется от актиновой нити, прикрепляясь к ней снова через такое же короткое время. Однако несмотря на чередование прикреплений и отделений, следующих с частотой от 5 до 50 раз в секунду, сила, развиваемая мышцей, остаётся неизменной, так как статистически в каждый момент времени в прикреплённом состоянии находится одно и то же количество мостиков [31].
Расслабление мышцы происходит после прекращения поступления двигательного импульса. При этом ионы кальция переходят обратно в цистерны саркоплазматического ретикулума. Уборка ионов кальция происходит в сторону с более высокой концентрацией этих ионов, поэтому этот процесс требует дополнительных затрат энергии. Процесс уборки ионов кальция называют кальциевым насосом и производится он за счёт энергии, получаемой при расщеплении АТФ, причём на уборку каждого иона кальция затрачивается две молекулы АТФ [24]. Снижение концентрации кальция в саркоплазме вызывает изменение пространственной ориентации тропонина, что в конечном итоге приводит к невозможности образования поперечных мостиков между толстыми и тонкими нитями. За счёт упругих сил, возникших ранее (при мышечном сокращении) в коллагеновых нитях, окружающих мышечное волокно, мышца расслабляется и возвращается в исходное положение, чему также может способствовать и сокращение мышц-антогонистов [11].
7.2.3 Изменение величины силы в фазе подъёма
Количество поперечных мостиков, связывающих актиновые и миозиновые нити, а, следовательно, и развиваемая мышечная сила согласно теории скользящих нитей, зависит от степени перекрытия толстых и тонких нитей, а значит, и от длины саркомера или мышцы. Максимум развиваемой изолированной мышцей силы будет наблюдаться при длине, примерно соответствующей состоянию покоя. При уменьшении длины сила уменьшается из-за того, что актиновые и миозиновые нити начинают мешать друг другу, а при растяжении мышцы до большей, чем в покое, длины сила уменьшается из-за того, что нити актина оказываются вытянутыми из миозиновых пучков. При этом только часть головок миозина может присоединиться к актину [31].
В реальных условиях мышцы, прикрепляющиеся к костям, вызывают движение рабочих звеньев тела. С движением рабочего звена меняется угол в суставе и, следовательно, длина обслуживающих данное сочленение мышц и угол подхода их к месту прикрепления на кости. При этом увеличиваются или уменьшаются плечо и момент силы мышц, что, в свою очередь, изменяет механические условия их работы. Эти условия могут быть выгодными, когда силовой потенциал используется полностью, и невыгодными, когда максимальное напряжение мышц используется только частично [18].
Сила, проявляемая в наименее целесообразной с механической точки зрения фазе движения, часто составляет не более 50-60% (рисунок 7.7) от силы в наиболее целесообразной его фазе [23].
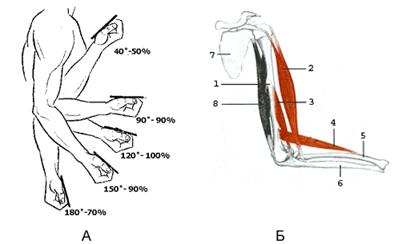
Рисунок 7.7 А - динамика максимальной силы при сгибании руки в локтевом суставе (по Платонову В.Н.).
Б - рисунок, иллюстрирующий сгибание руки в локтевом суставе (по [27]).
1 – плечевая кость; 2 - двуглавая мышца плеча; 3 - плечевая мышца; 4 - плечелучевая мышца; 5 – лучевая кость; 6 – локтевая кость; 8 - трёхглавая мышца плеча (разгибатель локтевого сустава)
При подтягивании на перекладине наименее благоприятным является верхний участок траектории движения в фазе подъёма, который недостаточно подготовленные спортсмены проходят с большим трудом, особенно в конечной части выполнения упражнения.
В начале подтягиваний, когда силовые способности спортсмена находятся на должном уровне, он может проходить проблемный участок по инерции – за счёт набранной ранее скорости. По мере накопления усталости силовые способности спортсмена снижаются настолько, что он уже не может набрать необходимую скорость. Поэтому в случае недостаточного уровня развития силы спортсмен «зависает» на верхнем участке траектории движения, затрачивая на его прохождение неоправданно большое количество энергии.
На рисунке 7.8 приведены графики зависимости резерва силовых способностей от высоты подъёма для 6 человек – 5 спортсменов (КМС и МС по полиатлону) и одного человека, не занимающегося спортом. Резервом силы в данном случае считается сила, которую способен развить испытуемый в какой-либо точке траектории движения сверх веса своего тела. Поскольку рост (а значит, и высота подъёма) и вес тела спортсменов различен, их силовые способности следует выражать в относительных единицах. Так, высота подъёма из исходного положения до уровня грифа принята за 100%, а резерв силы выражен в долях веса тела каждого из участников эксперимента.
В нижней части траектории движения (от 0 до 30 %) характер изменения резерва силы может быть различным, но, начиная примерно с 30% от высоты подъёма, резерв силы непрерывно снижается, что является экспериментальным подтверждением тезиса о проблемности верхнего участка траектории.
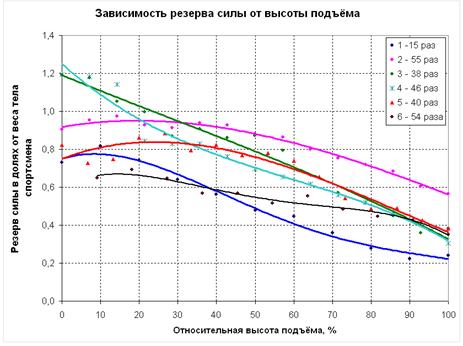
Рисунок 7.8 Изменение относительного резерва силы в фазе подъёма.
Взаимосвязь относительного резерва силы и результата в подтягивании имеет сложный характер. С одной стороны, испытуемый, не занимающийся подтягиванием (кривая 1), имеет более низкие показатели резерва силы в верхней части траектории движения по сравнению с квалифицированными полиатлонистами. Но с другой стороны, спортсмены, имеющие различные результаты в подтягивании (от 38 до 54 раз – кривые 3-6), показали в эксперименте практически равные значения относительного резерва силы на верхнем участке траектории движения. В то же время у спортсменов, имеющих практически равные результаты в подтягивании (54 и 55 раз – кривые 6 и 2), наблюдаются существенно отличающиеся показатели резерва силы на уровне грифа – соответственно 35% и 56% от веса тела. Это можно объяснить тем, что при достижении определённого уровня квалификации в подтягивании (предположительно 35-40 раз) первостепенное значение имеет не столько величина резерва силы, которая зависит от анаэробных возможностей мышц спортсмена, сколько скорость его расходования при выполнении подтягиваний, которая в большей степени определяется аэробными возможностями мышц.
Так, для спортсмена 2, резерв силы которого на уровне грифа перекладины составляет 56% от веса тела, средняя скорость расходования этого резерва будет составлять 56/55=1,02 % от веса тела в расчёте на одно подтягивание. Для спортсмена 6, резерв силы которого в той же точке составляет 35% от веса тела, скорость его снижения будет равна 35/54=0,65% от веса тела в расчёте на одно подтягивание. Следовательно, спортсмен 6, обладающий существенно меньшим резервом силы, выполняет подтягивания более экономично.
В параграфе 2.4.2 мы рассматривали соотношение между силой и скоростью мышечного сокращения (кривая Хилла), отметив, что скорость сокращения мышцы нелинейно снижается с увеличением нагрузки. Ненагруженная мышца сокращается с максимальной скоростью, но эта скорость зависит от типа мышечных волокон. Чем быстрее (при участии АТФ) происходит циклическое взаимодействие миозиновых мостиков с актином, тем выше скорость скольжения. АТФазная активность миозина в быстрых мышечных волокнах выше, чем в медленных, поэтому поперечные мостики волокон типа IIА и IIВ будут быстрее расщеплять АТФ и взаимодействовать с актином.
В условиях противодействия нагрузке максимальная скорость сокращения мышцы будет тем ниже, чем выше нагрузка. В случае если нагрузка не превышает предельные возможности спортсмена, он может произвольно менять скорость мышечного сокращения (например, скорость в фазе подъёма туловища). Так, когда все мышечные волокна участвуют в поднимании груза, относительная нагрузка на каждое мышечное волокно меньше (а скорость их сокращения больше), чем в том случае, когда активна лишь часть волокон. Таким образом, можно увеличить скорость укорочения мышцы при одной и той же нагрузке за счёт вовлечения дополнительных двигательных единиц [31].
Теоретически увеличение максимальной произвольной силы спортсмена должно приводить к улучшению результата в подтягивании за счёт относительного снижения нагрузки на мышцы и возможности подключения дополнительных двигательных единиц в ходе выполнения подтягиваний. Но на практике это часто происходит только в том случае, если длительность подтягиваний не превышает 1-2 минуты. И связано это с тем, что увеличение максимальной силы требует проведения соответствующих тренировок с большими отягощениями, которые направлены на увеличение количества миофибрилл и повышение концентрации креатинфосфата в быстрых мышечных волокнах. Но тренировки, состоящие из подходов с большими отягощениями, отрицательно сказываются на длительности виса. Получается замкнутый круг – увеличение одной способности происходит за счёт снижения другой, разорвать который можно только за счёт разумного сочетания тренировок, направленных на увеличение количества миофибрилл и тренировок, направленных на повышение числа митохондрий в рабочих мышцах спортсмена.
Дата публикования: 2014-10-20; Прочитано: 2549 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
