 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Режимы поиска цели в подсознании
|
|
Для нахождения веполяв режиме поиска в подсознании (назовем его поисковым веполем или ПВ) сначала определим расположение этого режима на оси времени относительно сознательных режимов. С большой долей уверенности можно утверждать, что поиск в подсознании начинается позже, чем поиск в сознании. Например, при решении задачи по АРИЗу сначала изобретательской ситуацией загружается сознание. Поэтому будем считать, что исходной моделью для задачи достройки поискового веполя является неполный веполь (рис.2.1в). Окончание поиска прекращается в момент захвата цели, когда Х-элемент из подсознания пробивается в сознание (веполь захвата на рис. 2.2а). Следовательно, поисковый веполь должен располагаться между неполным веполем и веполем захвата.
Предположим, что переход поиска из сознания в подсознание является некоторым пограничным режимом, имеющим черты как сознательного, так и подсознательного. Тогда этот пограничный режим должен иметь соответствующее вепольно-математическое представление. Пусть общей чертой с сознательным режимом будет математическое описание, т.е. система уравнений (2.1)-(2.2), а различием будет их структурное представление. Другая структура получается путем свертывания неполного веполя (рис.2.1в) по линии би-моно, т.е. от двух блоков умножения - к одному (рис.2.1г). Назовем вновь полученную структуру бисверткой. Возможность свертывания определяется избыточностью системы уравнений (2.1)-(2.2). Действительно, для задания положительной и отрицательной S-кривых достаточно дифференциального уравнения для одной координаты, а другая получается как зеркальное отображение первой. Избыточность гомеостаза является необходимым условием саморазвития.
В теории популяций блок умножения отвечает за передачу наследственной информации, за способность к воспроизводству, так как произведение xy характеризует рождаемость и равно числу попарных встреч особей мужского и женского полов популяции. Таким образом, в структуре бисвертки объединяется наследственный аппарат противоположных свойств технического противоречия, поэтому структурно бисвертка может быть названа веполем зачатия Х-элемента (или информационного поля нового решения). Закономерен вопрос: что именно является носителем наследственной информации ТП? Наиболее подходящим кандидатом на эту роль является физическое противоречие, т.е. можно предположить, что наследственными признаками ТП, передаваемыми Х-элементу, являются противоположные свойства физического противоречия. Поскольку ТП формулируется обычно на шагах 1.2 и 1.3 АРИЗ 85В, а физическое противоречие гораздо позднее, бисвертка передает наследственные признаки скрытно, т.е. в подсознании. После этого собственно и начинается поиск Х-элемента в подсознании.
Для получения структуры и математической модели поискового веполя используем единую форму записи систем дифференциальных уравнений (2.1)-(2.2) и (2.3)-(2.5). Любое из уравнений, например, для координаты x, можно записать в виде
dx / dt =c 1 xy + c 2 x+ c 3 y + c 4 z. (2.7)
Действительно, назначая c 1=-3/ K, c 3=- a / K, c 2= c 4=0, получаем уравнение (2.1). Назначая c 1=-3/ K, c 4= a / K, c 2= c 3=0, получаем уравнение (2.3).
Рассмотрим влияние слагаемых в правой части уравнения (2.7) на координату x. Как уже отмечалось выше, слагаемое c 1 xy отвечает за передачу наследственной информации, за способность к воспроизводству координаты x. Слагаемые с "чужими" координатами, т.е. c 3 y и c 4 z, определяют некоторое принуждение для движения координаты x, напротив, слагаемое c 2 x со "своей" координатой задает свободное развитие x.
Разглядывая структуру бисвертки (рис.2.1г), замечаем, что она полностью симметрична относительно продольной оси. Однако, с точки зрения математики [29], необходимым условием самоорганизации является снижение степени симметрии, ее потеря. Например, в физике жидкость считается симметричной, т.е. однородной и изотропной по всем направлениям. После замерзания в ней появляются выделенные направления, определяемые кристаллической решеткой, следовательно, степень симметричности снижается. Поэтому будем нарушать симметрию бисвертки, используя биологическую аналогию, тем более, что мышление относится к биологическим процессам.
Выше противоположные свойства ТП были определены как "родители" Х-элемента. Поэтому условно назовем кружок x "папой", кружок y - "мамой", а Х-элемент - "бэби". Какова же предпосылка разделения ранее равнозначных свойств ТП? Это - шаг 1.4 АРИЗа, на котором выбирается одна из схем конфликта: ТП-1 или ТП-2, обеспечивающая главную производственную функцию. Выбор позволяет:
нарушить симметрию бисвертки и поставить полезные свойства ТП в неравнозначное положение при дальнейшем поиске решения, обнаружить цель - прообраз Х-элемента.
Обнаружение в системах слежения за подвижной целью есть определение некоторого признака цели. В данном случае этим признаком является знак Х-элемента, так как знак задает место Х-элемента относительно того или иного свойства технического противоречия, а, если проще, то место, где он «спрятался» или куда мы его прячем в нашем подсознании. Это место далее, на шаге 1.6, задает движение Х-элемента относительно "родителей": кому он должен не мешать выполняться и вредное действие кого он должен устранять. Но пока он еще не вступил во взаимодействия с "родителями", то обладает только собственным свободным движением, что отражается стрелкой z с самого на себя (рис.2.5а).

Рис. 2.5. a) Структура СМС в режиме обнаружения Х-элемента; б) Структура СМС в режиме распознавания цели; в)поисковый веполь (ВП)
Пусть главную производственную функцию обеспечивает то полезное свойство ТП, которое определяется координатой x. Поэтому на шаге 1.4 делается выбор в пользу "папы". Тогда можно считать (с учетом следующего шага 1.5 по усилению x), что одна сторона конфликта, связанная с "папой", разрешена. "Папа" сделал свое дело, "папа" может пока свободно гулять. Для такой возможности предоставляем "папе" некоторую свободу в виде добавки собственного движения по полукруглой стрелке x (рис. 2.5б).
Теперь становится ясно, кому не должно мешать "бэби", а кого устранять. "Бэби" не должно мешать "папе", а должно вступить в борьбу (гомеостаз) с "мамой", поэтому проводим стрелку z от "бэби" к "маме". "Мама" временно теряет способность к воспроизводству, так как начинает выращивать "бэби", поэтому разрываем связь xy от общего блока умножения к "маме" и проводим стрелку y от "мамы" к "бэби". Получаем структуру СМС в режиме распознавания цели (рис.2.5б) и поисковый веполь - ВП (рис.2.5в). Основываясь на полученной структуре, запишем систему дифференциальных уравнений, описывающих ПВ,
dx / dt = c 1 xy + c 2 x+ c 3 y, (2.7)
dy / dt = c 4 x + c 5 z, (2.8)
dz / dt = c 6 y + c 7 z. (2.9)
Выбирая следующие значения коэффициентов: c1 = c6 = - c4 = -c5 = 1, c2=-4.5, c3=0.3, c7=0.38, получаем уравнения странного аттрактора Рёсслера[30], демонстрирующего винтовой хаос (рис. 2.6а).
Траектории изображающей точки аттрактора в трехмерном пространстве координат x, y, z начинаются по обе стороны от стационарной неустойчивой точки x =0, y =0, z =0 типа "седло-фокус". Движения в аттракторе Рёсслера приближенно можно разделить на два типа, переходящих друг в друга: раскручивающиеся спирали на "лепестке" y ↔ z и уход и возврат на "лепесток" по координате x, причем точки ухода и возврата на спирали являются хаотичными. Приближенность заключается в том, что "лепесток" y ↔ z не лежит в плоскости x =0, так как траектория иногда проскакивает ниже плоскости x =0, хотя подавляющая часть движений по координате x положительная. Очевидно, что эти два типа движений определяются двумя гомеостатами: спирали на "лепестке" y ↔ z - это гомеостат "мама" - "бэби", а уход и возврат - гомеостат "папа" - "мама" (x ↔ y).
Обратим внимание, что аттрактор Ресслера несимметричен как по вепольной структуре (рис.2.5в), так и траекторно (рис.2.6а). Центром "притяжения" и "отталкивания" в аттракторе является так называемая гомоклиническая орбита (рис.2.6б), замкнутая трехмерная кривая, проходящая через точку "седло-фокус" в начале координат. Так как гомоклиническая орбита замкнута, то движение изображающей точки по ней будет не хаотическим, а регулярным колебанием. Однако такое движение неустойчиво, как например, движение шарика вдоль верхушки хребта. Физически попасть на хребет можно (сверху), но двигаться вдоль него нельзя, так как _ иному склонам. Математически эта ситуация выражается в том, что для гомоклинической орбиты коэффициенты ci уравнений (2.7)-(2.9) являются числами в виде бесконечной непериодической дроби, реализовать которые при компьютерном моделировании невозможно. Следовательно, гомоклиническое колебание представляет собой идеальный конечный результат (ИКР), который может быть назван динамическим ИКР. Динамический ИКР представляет собой попытку подсознания одновременно реализовать два устойчивых состояния равновесия, соответствующие двум противоположным максимально полезным свойствам ТП y max и x min (рис.2.1а). Однако в пространстве они разнесены по разным сторонам прототипа, поэтому попытка осуществляется во времени.
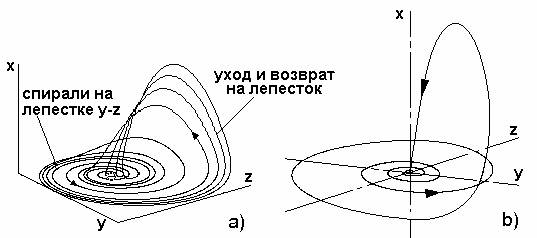
Рис. 2.6. а) Трехмерный аттрактор Рёсслера; б) гомоклиническая орбита
Можно заключить, что динамический ИКР обладает противоречиями: физическим - попасть на ИКР возможно, но удержаться на нем нельзя; математическим - уравнение ИКР существуют, но его решение в рациональных числах не представимо; психологическим - мышление притягивается к ИКР из-за его идеальности, но отталкивается от него из-за невозможности реализации.
Графики решения уравнений (2.7)-(2.9) с начальными условиями y (0)=- x (0)=2 и z (0)=0 приведены на рис. 2.7а. Кривые x (t), y (t), z (t) представляют собой хаотические колебания, на первый взгляд, не имеющие никакой связи друг с другом. Однако это не так. Кривая z (t) ("бэби") старается, по мере возможности, следить за кривой y (t) ("мамой"), т.е. повторять ее колебания. Конечно, амплитуды и фазовые сдвиги той и другой кривых меняются, однако сохраняется равномерность чередования максимумов и минимумов во времени: сначала идет максимум y (t), затем максимум z (t), потом двойка минимумов y (t) и z (t), затем снова двойка максимумов y (t) и z (t) и т.д. Cогласованность подтверждается также тем, что на «лепестке» аттрактора координаты y (t) и z (t) образуют плавную спираль, по крайней мере, от одного возврата на «лепесток» до следующего ухода. Такое явление можно называть стохастическим резонансом или синхронизацией хаоса [31].
В то же время задача распознавания может рассматриваться и как задача формирования образа, в данном случае, выращивания образа Х-элемента. Из имеющихся в подсознании стереотипов разрешения конфликтов в прошлом, координата y (t), как параметр порядка в терминологии Г.Хакена [32], "втягивает" в слежение за собой наиболее подходящий стереотип движения z (t). Если он не совпадает с необходимым движением, то по цепи обратной связи происходит коррекция системы мысленного слежения, чтобы движения в СМС приближались к ИКР. Происходит это следующим образом.
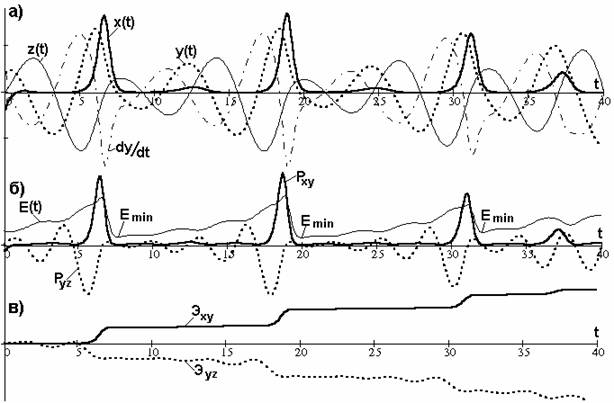
Рис. 2.7. а) координатные колебания; б)графики мощностей гомеостазов и ошибки СМС; в)энергетическая передача наследственной информации
Как видно из рисунка 2.7а, координата x (t) ("папа"- жирная сплошная линия) практически все время положительная, хотя при поиске в сознании (рис. 2.1а) она отрицательная. В этом инвертировании проявляется хорошо известное свойство отражения образов в мышлении. Каждый из импульсов x (t) представляет собой уход и возврат на "лепесток", а между импульсами x (t) происходит движение на "лепестке", характерное тем, что колебание координаты z (t) ("бэби"- тонкая сплошная линия) происходит примерно в противофазе с изменением скорости dy / dt (штрих-пунктирная линия) движения координаты y (t) (пунктирная линия). Следовательно, для движения на "лепестке" можно записать
dy / dt= - z (t) или 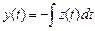 . (2.10)
. (2.10)
Формула (2.9) означает, что за время распознавания или интегрирования "бэби" выращивается таким, чтобы к моменту захвата интегральное значение "бэби" устраняло бы вредное действие "мамы". Как только колебание "бэби" отклоняется от предписанного уравнением (2.10), происходит коррекция всех движений в СМС путем подачи импульса x (t). По окончании коррекции СМС приходит в состояние с наибольшей вероятностью захвата цели или "озарения". На рис.2.7б эти состояния обозначены через E min, а график E (t) (тонкая сплошная линия) представляет изменение ошибки СМС. Ошибка определяется как эвклидова норма координат, т.е.
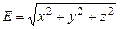 . (2.11)
. (2.11)
Равенство нулю ошибки (2.11) означает, что СМС попадает в начало гомоклинической орбиты аттрактора Рёсслера, т.е. в точку статического ИКР, когда противоречие полностью разрешено (x = y =0) без участия Х-элемента (z =0). Так как ИКР не может быть реализован, то захват происходит в точке E min локального (или глобального) минимума ошибки. Надежность захвата обеспечивается также нулевыми значениями мощности P xy как управляющего гомеостата, так и мощности P yz управляемого гомеостата в точках минимумов ошибки (рис.2.7б). Мощность гомеостаза переменных оценивается как величина их произведения, т.е. P xy = xy и P yz = yz и по сути является мощностью сигнала наследственной информации. Психологическая энергия сигналов наследственной информации для управляющего (Э xy) и управляемого (Э yz) гомеостатов вычисляется по формулам  а графики их изменения приведены на рис.2.7в. Как видно, управляемая энергия зеркально отражает управляющую энергию с небольшими искажениями.
а графики их изменения приведены на рис.2.7в. Как видно, управляемая энергия зеркально отражает управляющую энергию с небольшими искажениями.
Режим захвата демонстрирует рис. 2.8. При захвате структура СМС переходит в режим слежения со структурой, изображенной на рис.2.2 и уравнениями (2.3)-(2.5). Устойчивость СМС гарантирует сходящиеся процессы.
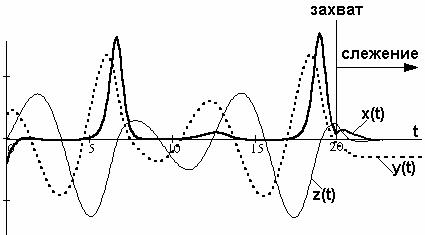
Рис. 2.8. Режим захвата и слежения
В результате анализа установлена вепольная структура и математическая модель системы слежения за новой идеей в мышлении изобретателя, включающая поиск, обнаружение, распознавание и захват образа Х-элемента.
Синтезирована математическая модель поисковых колебаний в виде хаотического аттрактора Ресслера, обладающая высоким поисковым потенциалом.
Определены понятия динамического и статического ИКР, а также получен математический критерий захвата Х-элемента (или "озарения") по минимуму отклонения системы слежения от ИКР.
Установлен скрытый механизм передачи Х-элементу наследственной информации (физического противоречия) от противоположных свойств технического противоречия. Механизм представляет сцепленные управляющий и управляемый гомеостаты, причем психологическая энергия управляемого гомеостата является зеркальным отражением психологической энергии управления.
Дата публикования: 2014-11-03; Прочитано: 452 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
