 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Глава 3. Неотения
|
|
Возможен еще один путь усиления интенсивности потребления кислорода у возникающей более прогрессивной группы животных - путь неотении, т.е. переход к активной самостоятельной жизни на ранних стадиях постэмбрионального, а иногда и поздних стадий эмбрионального развития (Зотин, Криволуцкий, 1982). Известно, что интенсивность дыхания на эмбриональных и личиночных стадиях развития животных значительно выше, чем во взрослом состоянии (Зотин, Зотина, 1993). Поэтому неотения приводит к появлению животных с более высоким уровнем стандартного обмена по сравнению с предковыми формами.
В истории сравнительной анатомии беспозвоночных на протяжении последних 150 лет было высказано немало предположений о происхождении отдельных крупных таксонов от преимагинальных стадий других групп животных. В случае насекомых, например, их происхождение от ранних стадий развития многоножек сомнений не вызывает. Этому соответствует и более низкий уровень интенсивности потребления кислорода взрослыми формами многоножек по сравнению с насекомыми: в классе Myriapoda a =0.45, Cr =5.78 (92), в то время как даже у низших насекомых отряда Colombota a =0.63, Cr =8.32 (97).
Считается, что не только насекомые и некоторые другие группы беспозвоночных возникли путем неотении, но и некоторые группы позвоночных (Воробьева, 1992; Иорданский, 1994) и многие группы растений (de Beer, 1958; Тахтаджан, 1970; Капанадзе, Капанадзе, 1995).
Явление неотении в случае измельчания вида или перехода его в неблагоприятные условия существования связано с трудностями борьбы за существование взрослых форм животных. Дальнейшая эволюционная судьба
неотенического вида, как и в случае измельчания, зависит от способности животных изменить коэффициенты в аллометрическом соотношении (46). В этом случае возможны три пути изменения энергетического обмена у возникающих групп животных (рис.38). Два из них, когда 0.75<k<1.00 и когда k=0.75 (если при этом возрастает коэффициент а), ведут по пути биоэнергетического прогресса, один, когда k<0.75, ведет в эволюционную ловушку. Неотенические виды имеют преимущество в энергетическом обмене при малых массах и недостаток - при больших по сравнению с предковыми формами.
В настоящее время существуют и процветают животные, которые не перешли полностью на путь неотении, но уже способны размножаться на личиночных стадиях развития. Другими словами, они могут существовать и давать потомство, не переходя во взрослое состояние. Стоит им потерять взрослые формы, и они превратятся в новый вид, взрослыми стадиями которого будут личинки предков.
Это, прежде всего, относится к семейству Ambystomidae из отряда хвостатых амфибий. Личинки этих амфибий - аксолотли - способны, как и взрослые формы - амбистомы - к размножению. Так как на личиночных стадиях развития интенсивность потребления кислорода выше, чем у взрослых форм, то к моменту половозрелости интенсивность дыхания аксолотлей выше, чем амбистом (Владимирова и др., 1993). Если бы представители рода Ambystoma полностью перешли на личиночный тип жизни, то образовавшийся новый вид обладал бы большей величиной коэффициента а и критерия упорядоченности.
Подтверждение этому выводу находим в работах Владимировой, Сербиновой (1992), Владимировой и др.(1993), в которых показано, что интенсивность потребления кислорода хвостатыми амфибиями в 2 раза ниже, чем бесхвостых (рис.24). В то же время, исследованный авторами вид Ambystoma mexicanum по уровню стандартного обмена занимают двоякое положение: у половозрелых амбистом, как показывает расчет по данным Владимировой и др.(1993), а = 0.22, в то время как у половозрелых аксолотлей а = 0.49 мВт (табл.31). Другими словами, амбистомы стоят в системе хвостатых
амфибий на первом месте, а половозрелые неотенические личинки - аксолотли - на последнем и не отличаются по стандартному обмену от некоторых бесхвостых амфибий (табл.35). То же свойственно и некоторым другим видам хвостатых амфибий, имеющих способных к размножению неотенических личинок (Bentley, Shield, 1973; Shield, Bentley, 1973; Ultsch, 1974; Bentley, 1975; Withers, Hillman, 1981). Интенсивность дыхания половозрелых личинок этих амфибий, как и в случае аксолотля, близка к интенсивности дыхания бесхвостых амфибий. Таким образом, в этих группах животных формируется новый таксон класса амфибий, который по уровню энергетического обмена приближается к отряду бесхвостых амфибий.
Интересные результаты были получены при изучении интенсивности потребления кислорода при метаморфозе аксолотлей в амбистомы (Владимирова и др., 1993). Метаморфоз аксолотлей вызывали на разных стадиях роста добавлением в воду тиреоидина в концентрации 1 г/мл, как рекомендует Воронцова и др. (1952). Потребление кислорода измеряли монометрическим методом Варбурга при 20°С в специально изготовленных сосудах цилиндрической формы. Была исследована интенсивность потребления кислорода аксолотлями от стадии вылупления (масса тела 0.01 г) до максимальной массы тела в 130г.
Как видно из рис.47 интенсивность дыхания аксолотлей непрерывно уменьшается во время развития. Можно выделить два периода изменения интенсивности дыхания: до 2 г и после, - когда уже возможен метаморфоз аксолотлей в амбистом. Поэтому, сравнивать дыхание аксолотлей и амбистом лучше во втором возрастном периоде. Половозрелые аксолотли имели массу тела в среднем равную 77 г и интенсивность потребления кислорода 0.202 мВт/г. Половозрелые амбистомы имели массу тела в среднем 34 г и интенсивность потребления кислорода 0.112 мВт/г
Аллометрическая зависимость скорости потребления кислорода от массы тела во втором возрастном периоде у аксолотлей имеет вид
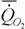 = 0.139· М 0.87 (181)
= 0.139· М 0.87 (181)
У амбистом на всех стадиях роста от 3 до 50 г интенсивность потребления кислорода была ниже, чем у растущих аксолотлей сходной массы (рис.47). Аллометрическая зависимость скорости дыхания от массы тела у амбистом имела вид:
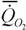 = 0.103 М 0.49 (182)
= 0.103 М 0.49 (182)
Из сопоставления этих формул видно, что коэффициент а у аксолотлей на 30% выше, чем у амбистом. Оказалось, что это справедливо и для межвидовой аллометрии, так как стандартный обмен аксолотлей разных видов примерно на 30% выше, чем у амбистом. Можно думать, поэтому, что увеличение коэффициента а и критерия упорядоченности у аксолотлей произошло за счет уменьшения коэффициента k [сравните аллометрические соотношения (181) и (182)]. А это означает, что только в том случае, когда коэффициент k для растущих личинок выше, чем у взрослых, неотения может привести к появлению видов с более высоким уровнем биоэнергетики. Возможно, что увеличение коэффициента а у насекомоядных млекопитающих (125) и колибриобразных птиц (127) произошло тем же путем, что и у аксолотлей. Не исключено, поэтому, что в основе образования указанных отрядов животных лежала неотения, связанная с переходом к взрослому существованию на ранних стадиях роста предков.
Дата публикования: 2014-11-04; Прочитано: 595 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
