 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Глава 1. Уменьшение размеров тела
|
|
Как отмечено выше, есть несколько путей преодоления неудачной ситуации, сложившейся в биоценозе, на пути биологического процветания той или иной группы организмов. Один из них связан не с пространственной миграцией этих организмов в места с не столь напряженной конкуренцией, а с уменьшением размеров тела. Можно себе представить, что представители некого вида животные или растения, не выдерживая конкуренции в борьбе за жизнь, постепенно мельчают, так как мелкие особи имеют шанс уцелеть в результате ли потери интереса к ним основных врагов, расширения ли их кормовой базы или возможности спрятаться при возникновении опасности. Проследим, какие же изменения могут произойти при уменьшении размеров организмов.
Прежде всего, уменьшение размеров тела приводит согласно формуле (47) к усилению интенсивности потребления кислорода. Если при этом меняющиеся организмы оказываются в такой нише биоценоза, где повышенный энергетический обмен дает решающее преимущество в борьбе за жизнь, то они прекращают процесс изменений в направлении уменьшения размеров и быстро увеличивают свою численность. Если же увеличение интенсивности
стандартного обмена не дает виду заметных преимуществ (не только энергетический обмен, но и многочисленные морфологические, функциональные и поведенческие особенности организмов определяют их конкурентоспособность), то вид продолжает изменение в сторону уменьшения размеров. В конце концов, может возникнуть ситуация, показанная на рис.44. На этом рисунке приведена зависимость интенсивности потребления кислорода от массы тела у млекопитающих. Кривая, соответствующая соотношению (47), которое для млекопитающих имеет вид (Зотин, 1988):
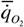 = 20.9М-0.26,
= 20.9М-0.26,
или (177)
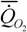 = 20.9М-0.74,
= 20.9М-0.74,
и носит название кривой Клайбера. Как видно из рис.26 она справедлива для всех млекопитающих, кроме насекомоядных [на схеме рис. 44 кривая (1)]. Расчет по данным, приводимым на рис.26, показал, что для насекомоядных аллометрическая зависимость определяется выражением:
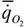 = 90,5М-0.60,
= 90,5М-0.60,
или (178)
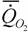 = 90.5М-0.40,
= 90.5М-0.40,
Теперь, на примере кривых, показанных на рис.44, продолжим рассмотрение гипотетического случая, когда под прессом естественного отбора происходит уменьшение размеров тела взрослых животных. По мере уменьшения массы тела в соответствии с кривой Клайбера [рис.44(2)] стандартный обмен млекопитающих возрастает. Это возрастание, начиная с массы тела порядка 20-30 г, может пойти в двух направлениях:
1) продолжить движение по кривой Клайбера [рис.44(2)],
2) вступить на более быстрый путь изменений [рис.44(1)].
В первом случае, если говорить об энергетическом обмене, ничего не изменится, так как сопоставимый стандартный обмен, т.е. коэффициент а из формулы (47), остается неизменным. Преимущества, которые при этом получают животные, связаны только с тем, что они занимают нишу, недоступную для других млекопитающих. Обладая повышенным по сравнению со всеми другими животными сходного размера (кроме птиц) стандартным обменом (рис.26) и связанными с этим возможностями, они могут найти свою нишу в биоценозе и прекратить дальнейший процесс уменьшения размеров. В смысле эволюционного прогресса уменьшение размеров тела в данном случае ничего не дает, так как критерий упорядоченности Сr, как и величина сопоставимого стандартного обмена, остаются прежними.
Во втором случае возможны два варианта.
Первый вариант. По мере уменьшения размеров организмов происходит изменение коэффициентов аллометрической зависимости (47) и стандартный обмен меняется по кривой 1, а не 2 (рис.44), т.е. возрастает значительно быстрее с уменьшением размеров тела чем по кривой Клайбера. При таком варианте животные получают значительные преимущества в величине энергетического обмена по сравнению с родительскими видами, а критерий упорядоченности у них намного больше, чем у всех остальных представителей данного класса. Для семейства землероек из отряда насекомоядных Сr=155 против Сr =104 (125) для всех остальных отрядов класса млекопитающих.
Сходные закономерности наблюдаются и среди класса птиц. Коэффициент к в аллометрической зависимости дыхания от массы тела птиц равен 0.71, в то время как в отряде калибриобразных 0.44 (126). Соответственно, в отряде калибриобразных Сr =286 против Cr =112 для всех остальных птиц (127).
Можно думать, что возникновение отряда насекомоядных в классе млекопитающих и отряда калибриобразных в классе птиц связано с изменением коэффициентов в аллометрическом соотношении (47). Это приводит к тому, что при уменьшении массы тела ниже 50-70 г стандартный обмен возрастает значительно быстрее, чем он должен был бы возрастать по кривой Клайбера, так
как аллометрическая кривая у насекомоядных и калибриобразных значительно круче, чем у остальных представителей классов, к которым относятся эти отряды [см., например, рис.44(2)]. Последнее определяет главный недостаток такого механизма прогрессивной эволюции: у крупных животных, вставших на указанный путь изменения энергетического обмена, несмотря на очень большую величину коэффициента а, а, следовательно, и критерия упорядоченности, стандартный обмен оказывается довольно низким. Расчет по формуле (178)

|
преимущество мелких насекомоядных в отношении величины стандартного обмена теряется по мере возникновения новых более крупных видов: начиная с массы в 70 г, стандартный обмен представителей этого отряда становится заметно меньше, чем у представителей других отрядов млекопитающих. Следовательно, повышенный энергетический обмен представителей нового таксона, возникшего в результате уменьшения величины коэффициента к в аллометрическом соотношении (46), ограничен областью мелких животных, т.е. имеется верхний предел размеров организмов для этого таксона. Снизу он также ограничен некоторой минимальной величиной, зависящей от возможности добычи и усвоения пищи. Дело в том, что повышенный энергетический обмен требует повышенного притока пищи. Известно, например, что мелкие землеройки из отряда насекомоядных потребляют за сутки количество пищи по массе больше, чем масса самого животного. Практически вся их жизнь состоит в непрерывном поиске пищи. Чтобы уменьшить суточную величину энергетических расходов они, подобно колибри и некоторым мелким рукокрылым, время от времени впадают в состояние оцепенения с очень низкой величиной энергетического обмена. Очевидно, что дальнейшее уменьшение
массы тела подобных животных может привести к полному прекращению активной жизнедеятельности. Для насекомоядных таким пределом является, по-видимому, масса тела меньше 2 г.
Таким образом, резкое возрастание энергетического обмена при уменьшении размеров тела в результате изменения коэффициентов в аллометрическом соотношении (46), (47) дает заметное преимущество видам, вставшим на этот путь арогенеза, но ограничивает их размеры довольно узкими рамками. Они попадают как бы в эволюционную ловушку (Зотин и др.,2000) и во всяком случае не способны вытеснить породившие их виды, имеющие массу тела выше верхнего предела ловушки.
Второй вариант. Это вариант арогенеза на начальных этапах совпадает с первым. Как и в последнем, по мере уменьшения размеров организмов происходит изменение аллометрической зависимости (47) и стандартный обмен меняется по кривой 1 [рис.44]. Это первый шаг на пути прогрессивной эволюции, позволяющий достигнуть величины сопоставимого стандартного обмена значительно большей, чем у породивших их видов. Происходит он в результате увеличения коэффициента b в соотношении (47). Второй шаг состоит в возвращении величины коэффициента b к прежнему значению при одновременном сохранении повышенной величины коэффициента а. В этом случае первоначальная кривая зависимости интенсивности дыхания от массы тела [рис.44(2)] должна сместиться вверх и все новые виды животных, независимо от их массы, будут иметь сопоставимый стандартный обмен выше, чем предковые виды. Возможным примером такого пути прогрессивной эволюции является класс птиц (рис.27).
Хорошо известно, что воробьиные птицы имеют стандартный обмен заметно выше, чем неворобьиные (Calder, 1984; Gavrilov, Dolnik, 1985; Шмидт-Ниельсен, 1987; Зотин, 1988). Расчет методом наименьших квадратов по данным, приводимым в обзоре В.М.Гаврилова и В.Р.Дольника (Gavrilov, Dolnik, 1985), показал (Зотин, 1988), что аллометрическая зависимость интенсивности потребления кислорода от массы тела у воробьиных птиц равна
 = 37.2M-0.28, (179)
= 37.2M-0.28, (179)
а у неворобьиных
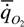 = 30.7М-0.29, (180)
= 30.7М-0.29, (180)
Соответственно, критерий упорядоченности у воробьиных птиц, согласно формуле (127), равен Сr=116.4, а у неворобьиных птиц - Сr=107.5. Так как коэффициент b y этих двух групп птиц практически одинаков, то стандартный обмен воробьиных птиц независимо от их размера всегда будет выше, чем стандартный обмен неворобьиных птиц той же массы. Это наглядно видно из рис.27, где приведены данные о стандартном обмене птиц разной массы и схематически показано на рис.44.
Некоторые авторы выделяют отряд воробьиных в отдельный подкласс, так как по численности видов, родов и семейств воробьиные превосходят всех остальных птиц вместе взятых. Это явно процветающая группа птиц наглядно демонстрирует преимущество повышения энергетического обмена и возможность на этой основе возникновения нового крупного таксона порядка отряда или даже класса.
Подводя итоги, можно сказать, что одноступенчатый механизм прогрессивной эволюции, возникающий при уменьшении массы тела животных, связанный с изменением коэффициента b в аллометрической зависимости (47), приводит к макроэволюционным изменениям животных в пределах эволюционной ловушки. Очевидно, что эволюционная ловушка может образовываться не только в процессе увеличения энергетического обмена при уменьшении размеров организмов, но и в результате чрезмерной специализации отдельных органов животных и растений, при переходе к паразитическому образу жизни и т.д.
Двухступенчатый механизм прогрессивной эволюции, первая ступень которого состоит в увеличении коэффициента b, a вторая - в стабилизации коэффициента а на высоком уровне при одновременном возвращении коэффициента b к начальному значению, приводит к макроэволюционным процессам образования крупного таксона животных или растений, неограниченных по размерам и эволюционным возможностям. Этот новый таксон обладает значительными
преимуществами в отношении энергетического обмена и способен вытеснить или, вернее, потеснить предковые формы во всех звеньях биоценоза.
Дата публикования: 2014-11-04; Прочитано: 747 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
