 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Автоволновые процессы в сердечной мышце
|
|
Мышечная ткань представляет собой возбудимую среду. Возбуждение, сообщаемое сетью нервных волокон, вызывает механохимические процессы. Процессы в возбудимых средах имеют важнейшее значение в физиологии, в частности в физиологии сердечной мышцы.
В возбудимой среде распространение импульса происходит без затухания за счет энергии клеток. Импульс генерируется каждой точкой среды. Пример однородной возбудимой среды - нервное волокно. Каждая точка возбудимой среды может находиться в одном из трех состояний: покоя, возбуждения, рефрактерности. Если на малый участок волокна, находящегося в состоянии покоя, подать возбуждение, то он перейдет в возбужденное состояние, в котором будет находиться некоторое время t, затем перейдет в рефрактерное состояние, в которое будет пребывать tR. В период рефрактерности tR участок не реагирует на возбуждающий сигнал, поэтому данный участок не может возбуждаться с частотой, большей t -1R.
В возбудимых средах реализуются автоволновые процессы. Источники волн могут возникать и в таких средах, в которых ни один из элементов не способен возбуждаться самопроизвольно, т.е. не является автоколебательным. Возникновение таких источников связано с особыми свойствами среды и волны возбуждения.
При рассмотрении реакций Белоусова - Жеботинского было отмечено существование источников волн двух типов - с ведущими центрами (ЭХО), посылающими концентрические волны, и с ревербераторами - вращаю-щимися спиральными волнами. Возникновение и последующее размножение ревербераторов играют важнейшую роль в механизме фибрилляции - очень опасной сердечной аритмии. Аритмия означает нарушение синхронизации распространения волн в сердечной мышце: вместо синхронных сокращений возникают беспорядочные колыхания, и организм быстро погибает, если не провести дефибриляцию.
Ревербератор в однородной двухмерной системе возникает, например, при наличии отверстия, периметр которого превышает длину волны l =Vt, где V – скорость распространения волны.
Циркуляция волны возбуждения (ревербератор) возможна и в непрерывной среде, неоднородной по рефрактерности. Если в некоторую область среды, имеющую форму кольца, послать импульс, то он будет циркулировать, так как волны возбуждения, распространяющиеся по часовой стрелке и против нее, встретятся и погасят друг друга. Если рефрактерность не всюду одинакова, то циркуляция возникает вследствие трансформации ритма (изменение ритма).
14. МЕХАНИЧЕСКИЕ СВОЙСТВА МЕМБРАН
14.1. Подвижность молекулярных компонентов в мембране
Гидрофобный эффект, объединяющий молекулярные компоненты в мембранах, препятствует их выходу в водную фазу за пределы мембраны. Силы мемжмолекулярного взаимодействия не мешают молекулам в мембранах обмениваться друг с другом местами, т.к. площадь контакта между водой и гидрофобными участками молекул при этом практически не изменяется. Таким образом молекулярные компоненты в мембранах сохраняют свою подвижность и могут диффундировать в пределах мембраны.
Молекулы липидов могут совершать вращательное движение вокруг своей длинной оси. Время корреляции вращательного движения tс молекул (время поворота на угол, равны 1 рад.) спин-меченых фосфолипидных молекул, стеаринов и жирных кислот в различных модельных и природных мембранах, находящихся в жидком состоянии, составляет примерно 10-9 с.
Вращательная подвижность сохраняется сравнительно высокой и при температуре ниже точки плавления жирнокислотных цепей липидов в мембранах.
Диффузионное перемещение молекул липидов вдоль слоя, называют латеральной диффузией. Oна происходит тоже довольно быстро (табл. 2). Коэффициент латеральной диффузии спин-меченных и флюоресцентных фосфолипидных зондов во многих искусственных и природных биологических мембранах составляет D = 10-7 - 10-8 см2/с. Среднеквадра-тичное расстояние, которое молекула проходит вдоль мембраны за время t, равно
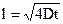
Легко вычислить, что при В= 6 10-8 см2/с молекула липида за 1 с перемещается по мембране на расстояние примерно 5000 нм, т.е. может пройти всю плазмaтическую мембрану таких клеток, как эритроцитов. Скорость латеральной диффузии существенно зависит от липидного состава мембран и температуры.
Энергия активации латеральной диффузии в липосомах из яичного лицитина и мембранах саркоплазматического ретикулума при комнатной температуре составляет 29 - 46 кДж/моль. При температурах ниже точки плавления углеводородных цепей липида константа латеральной диффузии снижается примерно на порядок.
Другой тип движения молекул липидов в мембранных системах - это трансбислойное движение (флип-флоп-переход). Оно происходит с относительно низкой скоростью вследствие высокого барьера для пересечения полярной головкой молекулы липида углеводородной зоны мембран. Скорость флип-флоп-перехода, оцененная по времени переноса половины количества молекул-меток из одного полуслоя в другой для модельных мембран, составляла 10 - 20 часов, для мембран эритроцитов - 20 - 30 мин., а для мембран электрических органов угря 3 - 7 мин.
Константы латеральной диффузии липидов
в искусственных и природных мембранах
| Мембраны | D, 108 см2 /с | Температура, оС | Метод |
| Яичный лицетин | 1,8 1,8 | ЯМР ЭПР | |
| Яичный лицетин + холестерин (4:1) Е.coli | 1,8 | ЭПР ЭПР ЯМР | |
| Седалищный нерв | 0,5 | ЭПР | |
| Липиды мембраны седалищно-го нерва | 0,8 | ЯМР | |
| Саркоплазматический ретикулум | 1,8 | ЯМР | |
| Липиды саркоплазматического ретикулума | 0,4 | ЯМР | |
| Микросомы печени | ЭПР |
Белковые молекулы в мембранах также проявляют диффузионную подвижность. Однако вследствие большей массы они движутся намного медленнее. Время корреляции вращения родопсина в фоторецепторных мембранах на три порядка меньше, чем для фосфолипидов.
Латеральная подвижность белков также ниже, чем у липидов. Коэффициенты латеральной диффузии белков в природных мембранах обычно находятся в пределах от 10-10 – 10-12 см2/с.
Таким образом, при физиологических условиях мембранные системы проявляют свойства двухмерной жидкости (исходя из коэффициентов для вращательной и латеральной диффузии). Вязкость таких сред формально определяется на основании уравнения Стокса-Эйнштейна:
h = k T/(6 p Dr),
где r – радиус диффундирующей молекулы.
Мембраны являются неоднородными и асимметричными, скорость вращения или латеральной диффузии определяется окружением молекулы и ее взаимодействием с другими молекулами. Поэтому вязкость, вычисленную из параметров подвижности того или другого зонда в мембране, рассматривают как "микровязкость". Это вязкость однородного эквивалента мембраны, в котором зонд проявляет аналогичную подвижность. Понятно, что величина вязкости будет зависеть от метода измерения.
14.2. Упругие свойства мембран
Под упругостью мембран понимают их способность изменять свое натяжение при растяжении-сжатии. Реакции мембран на внешние воздействия достаточно сложны и в настоящее время описываются только эмпирическими уравнениями. Однако при небольших амплитудах и низких скоростях деформации мембран под воздействием внешних сил можно рассматривать как термодинамически обратимые.
В равновесных условиях при постоянной температуре упругая потенциальная энергия мембран совпадает со свободной энергией Гельмгольца:
F = U – TS (14.1)
Упругий потенциал F при постоянной температуре равен работе W, совершенной над системой:
dF = dU – TdS = dW. (14.2)
Допущение, что мембрана изотропна вдоль поверхности, позволяет представить плотность потенциальной энергии упругости как функцию двух независимых переменных, характеризующих два типа деформации: относительного изменения площади поверхности a за счет изотропного растяжения-сжатия и параметра b, отражающего деформационное растяжение-сдвиг поверхности при постоянной площади. Например, растяжение круглой области в элипсоидную. Упругий потенциал можно представить через частные производные по каждому из этих параметров:
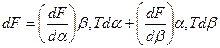 (14.3)
(14.3)
Противодействие изотропному растяжению-сжатию является свой-ством, характерным для твердого и жидкого состояний. В мембранах изотропное растяжение-сжатие площади поверхности происходит отно-сительно равновесного состояния. Это начальное изотропное натяжение Т в мембране можно оценить через частную производную свободной энергии в исходном состоянии (a = 0):
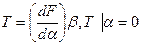 (14.4)
(14.4)
Связь изменения изотропного натяжения с малыми относительными изменениями площади (по сравнению с начальным состоянием) задается модулем поверхностного изотермического сжатия – модулем Юнга:
 |
При деформационном растяжении (при постоянстве площади) накоп-ление энергии и сопротивление деформации характеризуются поверх-ностным модулем упругости мембран при сдвиге m. Этот модуль опреде-ляется как частная производная упругой энергии деформации по параметру деформации b при постоянной площади и температуре:
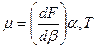
Эти два модуля являются важнейшими характеристиками мембран при механических воздействиях. Следует отметить, что упругие свойства природных мембран зависят от наличия в них белковых молекул. Модель-ные расчеты и непосредственные измерения показали, что величина поверхностного модуля упругого сжатия мембран эритроцитов составляют 3 – 4 10-3 Н/см, а поверхностный модуль сдвига равен примерно 10-5 Н/см. Из этого следует, что природные мембраны сильно сопротивляются изменению поверхностной плотности или площади, но сравнительно легко деформируются без изменения площади мембраны.
Дата публикования: 2015-11-01; Прочитано: 693 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
