 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Этап выдоха. 3 страница
|
|
ЭЛЕМЕНТЫ БИОФИЗИКИ ЗРЕНИЯ
СВЕТОПРЕЛОМЛЯЮЩАЯ СИСТЕМА ГЛАЗА
Органом зрения принято называть глаз. Однако, в зрительную сенсорную систему (зрительный анализатор) вместе с фоторецепторами (палочками и колбочками) и рядом нейронов, которые находятся в сетчатой оболочке глаза входят определенные отделы ЦНС. В то же время, глаз включает не только фоторецепторы и нейроны, но также разнообразные вспомогательные элементы зрительной рецепции. Эти вспомогательные элементы сами не воспринимают свет и не участвуют в переработке (передаче) сигналов от фоторецепторов, но способствуют восприятию света. Важнейшим вспомогательным аппаратом фоторецепции является диоптрическая светопреломляющая система глаза. Упрощенная схема разреза глаза человека имеет вид:

Схема горизонтального разреза глаза человека
1 – роговица; 2 – хрусталик; 3 – стекловидное тело, камерная влага;
4 – сетчатка; 5 – центральная ямка желтого пятна; 6 – зрительный нерв;
7 – сосудистая оболочка; 8 – склера; OO’ – главная оптическая ось;
KK’ -зрительная ось;  - угол между OO’ и KK’.
- угол между OO’ и KK’.
Светопреломляющая система глаза фокусирует изображение окружающего мира на сетчатой оболочке (сетчатке), в которой расположены фоторецепторы. Диоптрический аппарат глаза подобен сильной собирающей линзе, и, при рассмотрении далеких предметов, сводит в точку на сетчатке входящие в глаз параллельные лучи, то есть, действует наподобие объектива зрительной трубы. Фокусированные изображения осуществляются за счет преломляющей системы глаза, к которой относятся:
- роговица;
- хрусталик;
- камерная плата;
- стекловидное тело.
Преломляя световые лучи, они обеспечивают формирование на сетчатке действительного уменьшенного и обратного (перевернутого) изображения рассматриваемого предмета. Отрезок прямой, проходящий через геометрические центры роговицы и хрусталика, называют главной оптической осью глаза (ОО’). Она объединяет весь его диоптрический аппарат в центрированную светопреломляющую систему. Центрированной называют такую систему светопреломляющих поверхностей, геометрические центры которых лежат на одной прямой. Обратимся к идеальной центрированной системе Гаусса и попытаемся применить ее к человеческому глазу.
Центрированная диоптрическая система при построении изображения предметов рассматривается, как одно целое, и ход лучей в ней определяется при помощи шести, так называемых кардинальных точек, которые лежат на оси ОО’. К этим точкам относятся два главных фокуса (передний F1 и задний F2), две главные точки (Н1 и Н2) и две узловые точки (N1 и N2).

Схема построения оптического изображения
Главными фокуcами называют точки (F1 и F2), в которых после преломления собираются лучи, входящие в систему параллелей, так как коэффициенты преломления сред (n), находящихся перед глазом  , и внутри глаза (n=1,336), не одинаковы, то и фокусные расстояния
, и внутри глаза (n=1,336), не одинаковы, то и фокусные расстояния  . Размеры изображений предметов, построенных на h1 и h2, одинаковы. В местах пересечения главных плоскостей с главной осью расположены главные точки Н1 и Н2, от этих точек отсчитываются фокусные расстояния: от Н1 – переднее, f1, от Н2 – заднее f2.
. Размеры изображений предметов, построенных на h1 и h2, одинаковы. В местах пересечения главных плоскостей с главной осью расположены главные точки Н1 и Н2, от этих точек отсчитываются фокусные расстояния: от Н1 – переднее, f1, от Н2 – заднее f2.
Среди лучей, исходящих от предмета, всегда находится такой, который после преломления диоптрической системы, покидает ее без изменения первоначального направления. Этот луч претерпевает в системе только смещение, вследствие чего и распространяется по входу и выходу из нее, параллельно исходному направлению. Точки, в которых продолжение падающего луча и выходящего луча пересекают главную ось, называются узловыми точками. N1 – передняя узловая точка, N2 – задняя. Через эти точки падающий и преломленный лучи проходят под одинаковым углом по отношению к главной оси ( ), применительно к глазу, он называется углом зрения или зрительным углом.
), применительно к глазу, он называется углом зрения или зрительным углом.
ТОНКАЯ И ТОЛСТАЯ ЛИНЗЫ
Если вся центрированная преломляющая система находится в однородной среде (n1 = n2), то N1 совпадает с Н1, а N2 с Н2. Такой относительно простой центрированной диоптрической системой является тонкая линза, когда обе ее поверхности контактируют с воздухом. В тонкой линзе можно пренебречь ее толщиной по сравнению с расстоянием соответствующих фокусов от передней и задней поверхности, тогда, в этом случае, в центрированной системе точки N1 и N2, Н1 и Н2 являются центром оптической линзы, то есть сливаются в одну. При построении изображений, которые получаются в тонкой линзе, окруженной однородной средой, расстояние от оптического центра до предмета и расстояние до изображения от оптического центра, связаны с фокусным расстоянием по следующей формуле:
 - формула тонкой линзы.
- формула тонкой линзы.
 – расстояние от предмета до оптического центра;
– расстояние от предмета до оптического центра;
b – расстояние от оптического центра до изображения.
Для толстой линзы, чему можно уподобить центрированную систему глаза, формулы, связывающие и b, гораздо сложнее, они включают все кардинальные точки центрированной диоптрической системы. Для упрощения расчетов прибегают к моделям реальных диоптрических систем. При этом стремятся обойтись в модели минимальным количеством кардинальных точек.
ПРИВЕДЕННЫЙ РЕДУЦИРОВАННЫЙ ГЛАЗ
ЧЕЛОВЕКА
Моделью светопреломляющего аппарата органа зрения служит, так называемый, приведенный редуцированный глаз. Редукция означает упрощение, то есть сведение сложного к простому, более доступному для анализа. Физическим аналогом приведенного редуцированного глаза является стеклянная линза, которая одной поверхностью контактирует с воздухом, а другой – с жидкостью, обладающей n = 1,336. Первая поверхность направлена в пространство предметов, а другая поверхность линзы – к пространству изображений.
Диоптрический аппарат глаза не поддается точному математическому описанию. Это связано с тем, что глаза разных людей сильно различаются, а также из-за подвижности хрусталика, и по ряду других причин. Так, например, главная оптическая ось глаза человека проводится довольно приблизительно. Она не совпадает со зрительной осью КК’, которая проходит через геометрические центры роговицы и желтого пятна сетчатки. В направлении зрительной оси глаз имеет наилучшую разрешающую способность. Угол между главной оптической осью ОО’ и KK’, как правило не превышает 5 градусов. Он обозначается  обычно учитывается в офтальмологии при назначении очков.
обычно учитывается в офтальмологии при назначении очков.
Итак, человеческий глаз можно рассматривать в качестве центрированной преломляющей системы весьма условно. На рисунке приведена количественная характеристика одной из наиболее адекватных физических моделей человеческого глаза.

Глаз как центрированная светопреломляющая система
Из рисунка видно, что расстояние между Н1 и Н2 составляет примерно 0,25 мм. Столь близкое расстояние допускает замену двух главных плоскостей (h1 и h2) на одну главную плоскость. Точки N1 и N2 практически также совпадают в единую узловую точку глаза, ее называют оптическим центром глаза (между N1 и N2). В замене каждой из названных пар кардинальных точек на единые угловую и узловую и состоит смысл редукции центрированной диоптрической системы реального глаза. В схеме Гульстранда единая плоскость h и единая узловая точка N находятся на расстоянии 1,6 мм и 7,5 мм соответственно от передней поверхности роговицы. Все расстояния, кроме фокусных (f1, f2), отсчитываются от передней поверхности роговицы. По ходу лучей внутрь глаза эти расстояния имеют знак «+». В противоположном (наружном) направлении расстояния имеют знак «-».
Числа, характеризующие расстояние единых главной и узловой точек глаза, необходимы для расчета диоптрического аппарата человеческого глаза.
БИОФИЗИЧЕСКИЙ МЕХАНИЗМ
ВОСПРИЯТИЯ СВЕТА РЕЦЕПТОРАМИ
Свет, сфокусированный диоптрическим аппаратом глаза на сетчатке, возбуждает ее фоторецепторы. Он достигает фоторецепторов, пройдя всю толщу сетчатки, поскольку палочки и колбочки располагаются в тех слоях сетчатки, которые прилежат ближе к сосудистой оболочке. Свет проходит в сетчатке следующие слои, прежде чем достигнуть фоторецепторов:
1. Слой ганглеозных клеток.
2. Внутренний синоптический слой.
3. Внутренний ядерный слой, состоящий из амакриловых клеток, горизонтальных клеток (Г), биполярных нейронов (Б).
4. Наружный синоптический слой палочек и колбочек (П и К).
5. Внутренний ядерный слой П и К.
Каждый глаз человека состоит из примерно 125 млн палочек и примерно 5 млн колбочек.
Фоторецепторы П или К схематично можно представить в виде:

Схема строения фоторецепторной клетки (палочки или колбочки)
1 – наружная мембрана П или К; 2 – диски; 3 – митохондрии; 4 – аппарат Гольджи;
5 – ядро клетки; 6 – синоптическая зона: НС – наружный сегмент;
ВС – внутренний сегмент.
Диаметр П (К) составляет примерно 2 мкм, длина – примерно 50-60 мкм. Длина НС составляет примерно 20-30 мкм. Вся цитоплазма НС заполнена своеобразными органеллами, которые называют дисками. Промежутки между ними не превышают 15 нкм. В НС каждой П (К) содержится от 700 до 1000 дисков, которые напоминают собой сплющенные воздушные шары, сложенные в стопку. Диаметр каждого диска почти соответствует диаметру наружного слоя (примерно 2 мкм), а толщина диска составляет примерно 15-18 нм. Его внутреннее пространство представляет собой узкую щель, шириной = 1 нм. Эта щель расширяется до нескольких нм у краев диска. Таким образом, на внутридисковое содержимое вместе с цитозолем приходится не более половины объема НС, а другую половину занимают мембраны дисков.
НС буквально нафарширован мембранами, и именно в них осуществляется первичный процесс восприятия света. В этой связи, мембрану диска называют фоторецепторной мембраной.
Молекулярная организация фоторецепторной мембраны у позвоночных и беспозвоночных животных практически одинакова. Подобно всем БМ, она состоит из липидов и белков (протеинов), однако, ей присущи две кардинальные отличительные особенности. Во-первых, ее основным белковым компонентом является зрительный пигмент – хромогликопротеид родопсин. Его концентрация достигает 90 % от общего содержания протеинов в фоторецепторной мембране. Во-вторых, более 40 % жирокислотного состава фосфолипидов, образующих дисковые мембраны, приходится на полиненасыщенную декозагексоеновую кислоту, в состав которой входит 22 атома углерода с присутствием 6 двойных связей. Кроме нее, в состав мембранных фосфолипидов входят и другие полиненасыщенные длинноцепочечные жирные кислоты, содержащие от 4 до 6 двойных связей. Вместе они составляют более половины всех жирных кислотных остатков. Высокая степень ненасыщенности жирнокислотных «хвостов» фосфолипидных молекул придает гидрофобному бимолекулярному каркасу фоторецепторной мембраны необычайно высокую подвижность, вследствие чего данная мембрана обладает малой вязкостью по сравнению с другими мембранами.
РЕАКЦИЯ РОДОПСИНА НА ДЕЙСТВИЕ СВЕТА
Первичный механизм возбуждения П светом связан со сложными превращениями родопсина в фоторецепторной мембране. Родопсин – это высокомолекулярное соединение, состоящее из двух основных компонентов:
1. Альдегид витамина А (ретиналь).
2. Липопротеин под названием опсин (белок глаза).
Спектр поглощения родопсина имеет вид:

Спектр поглощения родопсина
Спектр поглощения родопсина хорошо приспособлен к восприятию света в достаточно широкой области солнечного излучения, но лучше всего этот зрительный пигмент поглощает свет в зеленой части этого спектра (500 нм). Красные и синие лучи он отражает, причем, в отраженном свете преобладает красный компонент, хотя присутствует и синий. Поэтому пигмент имеет пурпурный цвет, определивший и его название. Родопсин относится к наиболее интенсивно окрашиваемым соединениям из всех известных в настоящее время в органической химии. Возможность поглощения фотонов родопсина близка к теоретически возможному значению.
В темноте ретиналь пребывает в 11 цис-форме, для которой характерна некоторая скрученность молекулы. Скрученность объясняется тем, что в планарной форме между близко расположенными метильной группой у углерода (С13) и водородом у С10, возникает сильное отталкивание. Сила отталкивания не допускает плоского расположения  -связи. Скрученность в молекуле 11 цис-ретиналя обеспечивает наибольшую энергию взаимодействия с опсином, вследствие чего их комплекс (ретиналя и опсина) весьма устойчив. Для его разрушения необходимо преобразовать 11 цис-ретиналь в другой изомер, в так называемый полностью транс-ретиналь. Такое преобразование из цис- в транс-ретиналь происходит под действием света. Этот процесс называется фотоизомеризацией.
-связи. Скрученность в молекуле 11 цис-ретиналя обеспечивает наибольшую энергию взаимодействия с опсином, вследствие чего их комплекс (ретиналя и опсина) весьма устойчив. Для его разрушения необходимо преобразовать 11 цис-ретиналь в другой изомер, в так называемый полностью транс-ретиналь. Такое преобразование из цис- в транс-ретиналь происходит под действием света. Этот процесс называется фотоизомеризацией.
При поглощении фотона ( ), молекула родопсина претерпевает электрический переход «
), молекула родопсина претерпевает электрический переход « », при этом, молекула родопсина оказывается в синглетном возбужденном состоянии. Необходимо отметить, что в органических молекулах -орбитали, которые свойственны основному энергетическому состоянию, являются связывающими, то есть, они стремятся к перекрытию и образованию второй связи. Напротив,
», при этом, молекула родопсина оказывается в синглетном возбужденном состоянии. Необходимо отметить, что в органических молекулах -орбитали, которые свойственны основному энергетическому состоянию, являются связывающими, то есть, они стремятся к перекрытию и образованию второй связи. Напротив,  -орбитали, характерные для возбужденного состояния молекулы, относятся к разрыхляющим орбиталям. Их перекрытие энергетически невыгодно. В силу чего, части молекулы, соединенные двойной связью с участием -орбитали, разворачиваются на 90 градусов, и двойная связь превращается в одинарную. В этой связи, в молекуле ретиналя, возбужденной светом, происходит поворот атомов вокруг двойной связи. Такое измененние пространственного расположения атомов, входящих в состав ретиналя, требует затрат энергии. Эти энергозатраты характеризуются преодолением потенциального барьера. Для разворота атома вокруг двойной связи в 11 цис-ретинале достаточно энергии одного фотона. Для фотоизомеризации достаточно и времени, в течение которого молекула родопсина пребывает в сигнетном возбужденном состоянии (
-орбитали, характерные для возбужденного состояния молекулы, относятся к разрыхляющим орбиталям. Их перекрытие энергетически невыгодно. В силу чего, части молекулы, соединенные двойной связью с участием -орбитали, разворачиваются на 90 градусов, и двойная связь превращается в одинарную. В этой связи, в молекуле ретиналя, возбужденной светом, происходит поворот атомов вокруг двойной связи. Такое измененние пространственного расположения атомов, входящих в состав ретиналя, требует затрат энергии. Эти энергозатраты характеризуются преодолением потенциального барьера. Для разворота атома вокруг двойной связи в 11 цис-ретинале достаточно энергии одного фотона. Для фотоизомеризации достаточно и времени, в течение которого молекула родопсина пребывает в сигнетном возбужденном состоянии ( с), поскольку цис-тран-изомеризация ретиналя происходит всего за 0,1-0,3 пс, квантовый выход фотоизомеризации 11 цис-ретиналя в родопсине достигает 0,7.
с), поскольку цис-тран-изомеризация ретиналя происходит всего за 0,1-0,3 пс, квантовый выход фотоизомеризации 11 цис-ретиналя в родопсине достигает 0,7.
Фотоизомеризация – есть первая и единственная фотохимическая реакция в зрительном акте. Она протекает внутри отдельной молекулы родопсина и, в отличие от фотосинтеза, не сопровождается никакими химическими реакциями. Зрение отличается от фотосинтеза, который происходит в растениях, еще и тем, что между молекулами родопсина невозможен перенос энергии, так как, в фоторецепторной мембране молекулы родопсина удалены друг от друга на расстояние более 7 нм. Непосредственным следствием фотоизомеризации, приводящей к появлению у человека ощущения света, служат конформационные перестройки молекулы родопсина. Дело в том, что пространственно 11 цис-ретиналь точно соответствует конфигурации активного центра. При образовании транс-ретиналя, это соответствие (комплементарность) нарушается. В результате чего ретиналь отщепляется от опсина, и родопсин обесцвечивается, то есть, претерпевает фотолиз.
Вне действия света, распавшийся родопсин восстанавливается. В ходе восстановления, транс-ретиналь преобразуется в цис-форму. Такой переход обратной фотоизомеризации называется реизомеризацией. У позвоночных животных транс-ретиналь, отщепившись от опсина, поступает в пигментный эпителий сетчатки, где подвергается ферментативным превращениям. Из пигментного эпителия фоторецептором (П, К) доставляется 11 цис-ретиналь. Каждая палочка ежесекундно синтезирует  молекул родопсина из поступающего в нее 11 цис-ретиналя, который соединяется с опсином. Суточный синтез этого зрительного пигмента в целом оценивается величиной примерно
молекул родопсина из поступающего в нее 11 цис-ретиналя, который соединяется с опсином. Суточный синтез этого зрительного пигмента в целом оценивается величиной примерно  молекул.
молекул.
Конформационная перестройка родопсина, называющаяся фотоизомеризацией ретиналя, довольно существенна. Естественно, что перестройку сопровождают значительные изменения физико-химических свойств фоторецепторной мембраны, в которую встроен родопсин. Одним из важных последствий этих изменений служит сдвиг мембранного потенциала фоторецепторной клетки.
РЕЦЕПТОРНЫЕ ПОТЕНЦИАЛЫ
Сдвиг мембранного потенциала фоторецепторной клетки (П или К), который принято называть рецепторным потенциалом, происходит на ее плазмолемме, тогда, как фотоизомеризация родопсина совершается преимущественно во внутриклеточных мембранах, то есть, в мембранных дисках наружных сегментов.
Свойство фоторецепторных мембран изменяется одновременно с конформационной перестройкой в них опсина. После поглощения (кванта света), спустя 1,5-2 мс, наблюдается гиперполяризация плазмолеммы фоторецепторной клетки (П или К). Следовательно, в палочке (К) существует механизм передачи сигнала с фоторецепторных мембран дисков на плазмолемму П (К). В передаче участвует, так называемый, внутриклеточный посредник, природа которого до настоящего времени полностью не установлена, однако, считают, что в посредничестве участвуют циклический гуанозинмонофосфат (цГМФ) и ионы  .
.
В следствие конформационной перестройки молекул родопсина, в фоторецепторной мембране изменяется содержание внутриклеточного посредника в цитозоле наружного сегмента П или К. Это приводит к закрытию 100-300 натриевых каналов в плазмолемме, которые в отсутствие светового раздражителя пребывали в открытом состоянии. Они обладают свойствами потенциалнезависимых ионных каналов и, будучи управляемые светом, получили название фотозависимых натриевых каналов. Когда закрываются 100-300 каналов, электропроводимость плазмолеммы наружного сегмента падает на 1-3%. Падение мембранной электропроводимости приводит в гиперполяризации фоторецепторной клетки. Гиперполяризационный сдвиг мембранного потенциала П (К), возникающий под действием света, называют рецепторным потенциалом (РП). Принято считать, что ПП фоторецепторной клетки генерируется в темноте на плазмолемме внутреннего сегмента. По своей природе ПП является калиевым потенциалом, подобно всякому мембранному потенциалу в условиях покоя. В темноте ПП палочки (колбочки) составляет примерно от –20 до –40 мВ.
В системе генерации энергетического потенциала П плазмолемма наружного сегмента выполняет роль шунта с переменной электропроводностью. Электропроводность регулируется (управляется) светом при помощи фотозависимых натриевых каналов. Когда все натриевые каналы плазматической мембраны наружного сегмента одной П оказываются заблокированными светом, удельное поверхностное сопротивление этой мембраны достигает величины  . В темноте ионный ток, входящий через каналы в цитоплазму, довольно значителен, и составляет примерно 40 nА. А при освещении сетчатки он блокируется, при этом степень его ослабления зависит от интенсивности света. Чем слабее натриевый ток через плазматическую мембрану наружного сегмента, тем больше выражена гиперполяризация в фоторецепторной клетке. Изменение потенциала П (К) во времени имеет вид:
. В темноте ионный ток, входящий через каналы в цитоплазму, довольно значителен, и составляет примерно 40 nА. А при освещении сетчатки он блокируется, при этом степень его ослабления зависит от интенсивности света. Чем слабее натриевый ток через плазматическую мембрану наружного сегмента, тем больше выражена гиперполяризация в фоторецепторной клетке. Изменение потенциала П (К) во времени имеет вид:

Рецепторные потенциалы одиночной колбочки, возникающие в ответ
на короткие (10 мс) вспышки света при трех разных интенсивностях света
 – интенсивность света. Чем больше I, тем больше гиперполяризация
– интенсивность света. Чем больше I, тем больше гиперполяризация  .
.
При поглощении 1 фотона () гиперполяризация фоторецепторов разных представителей животного мира неодинакова. Так, в палочках сетчатки позвоночных животных гиперполяризация составляет десятки и сотни мкВ, тогда, как в фоторецепторных клетках членистоногих она достигает 10 мВ. Эти приведенные числа подтверждают положение об усилении сигнала в рецепторных аппаратах. Действительно, квант света, несущий энергию  Дж, вызывает фотолиз только одной молекулы родопсина, что в свою очередь приводит к закрытию в плазмолемме фоторецепторной клетки позвоночных животных 100-300 натриевых каналов. Это приводит к образованию РП. Однако, энергия РП на 3 порядка превосходит энергию вызвавшего его фотона.
Дж, вызывает фотолиз только одной молекулы родопсина, что в свою очередь приводит к закрытию в плазмолемме фоторецепторной клетки позвоночных животных 100-300 натриевых каналов. Это приводит к образованию РП. Однако, энергия РП на 3 порядка превосходит энергию вызвавшего его фотона.
Амплитуда рецепторного потенциала возрастает при повышении интенсивности света, падающего на сетчатку, причем, в пределах 1000-кратного изменения интенсивности, эта зависимость имеет логарифмический характер:
 - изменение амплитуды РП.
- изменение амплитуды РП.
 - коэффициент пропорциональности;
- коэффициент пропорциональности;
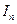 - интенсивность последующего;
- интенсивность последующего;
 - интенсивность предыдущего.
- интенсивность предыдущего.
При ярком освещении амплитуда рецепторного потенциала может достигать 25 мВ. Величина РП зависит также и от длины волны света. П или К в ответ на зеленый свет (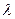 =500 нм), генерируют наибольший РП. Среди К выделено 3 типа:
=500 нм), генерируют наибольший РП. Среди К выделено 3 типа:
1) дает максимальный ответ РП на синий цвет;
2) дает максимальный ответ РП на зеленый цвет;
3) дает максимальный ответ РП на желто-красный цвет.
Использование микроэлектронной техники для изучения реакции фоторецепторов на облучение светом, позволило обнаружить деполяризацию П и К, при этом деполяризация предшествует гиперполяризации. Амплитуда деполяризации очень мала и составляет примерно 0,1 мкВ. Учитывая временные соотношения де- и гиперполяризации, первую из них стали называть ранним, а вторую – поздним РП, и обозначили, соответственно, РРП и ПРП. Установлено, что РРП обусловлен преимущественно перемещениями электрических диполей в мембранах наружного сегмента. Эти перемещения диполей связаны с фотолизом молекул родопсина. Другими словами, РРП обусловлен током смещения в фоторецепторных мембранах, и РРП не служит промежуточным звеном передачи сигналов от них к синоптической зоне П или К, при действии света. Эта роль принадлежит гиперполяризации. В отличие от других рецепторных аппаратов, где таким звеном служит деполяризационные сдвиги мембранного потенциала. Таким образом, только поздний РП является рецепторным потенциалом П или К в полном значении этого термина.
РАСПРЕДЕЛЕНИЕ СИГНАЛА ПО СЕТЧАТКЕ
Поздний РП, возникнув в НС, распределяется в синоптической зоне П или К по закону электротонической передачи (с декрементом). При этом, постоянная длины плазмолеммы фоторецепторной клетки, которая проводит гиперполяризацию, достигает 10 мкм. Достигнув синоптической зоны П (К) ПРП изменяет свойства ее мембраны и, тем самым, влияет на медиаторную систему, то есть, на передачу сигнала к биполярному нейрону. Химическая природа медиатора, выделяемого фоторецепторной клеткой, пока еще неизвестна точно, однако, наиболее вероятными кандидатами на эту роль признаются глутамат и аспарат. Обе аминокислоты обладают деполяризующим действием на постсинаптическую мембрану. При облучении П (К) светом, биполярные нейроны так же испытывают гипреполяризацию. Принято считать, что фоторецепторной клетке в темноте свойственен определенный темп выделения деполяризующего медиатора (глутамата или аспарата), а гиперполяризация синаптической зоны при освещении замедляет темп выброса медиатора, следовательно, постсиноптические мембраны, принадлежащие биполярным нейронам, также гиперполяризуются. Гиперполяризационный сдвиг мембранного потенциала биполярного нейрона электротонически распространяется по эго-плазмолемме к синапсу на теле третьего нейрона, который входит в состав афферентного пути сетчатки. Медиатор, поступающий от биполярного нейрона ганглиозной клетки, вызывает ее деполяризацию. Деполяризационный сдвиг потенциала распределяется электротонически с тела по аксонам ганглиозной клетки. Плазматическая мембрана аксона ганглиозной клетки (аксолемма) обладает возбудимостью, благодаря чему там и формируется ПД, когда деполяризация достигает порогового уровня. ПД, неся информацию от фоторецепторов, поступает в ЦНС по волокнам зрительного нерва. Эти волокна представляют собой аксоны ганглиозных клеток сетчатки. Схематически передачу поздних РП по сетчатке можно представить в виде:

Схема передачи фоторецепторного сигнала по афферентному пути в пределах сетчатки
РП – рецепторная палочка; НС – наружный сегмент; ВС – внутренний сегмент; СЗ – синоптическая зона; ГК – ганглиозная клетка; ПРП – поздний рецепторный потенциал; ПД – потенциал действия; ЦНС – центральная нервная система
Зрительный нерв включает примерно  волокон. По этим волокнам в мозг направляются сигналы от 130
волокон. По этим волокнам в мозг направляются сигналы от 130  фоторецепторов. Следовательно, в среднем 1 аксон ГК собирает информацию от множества рецепторных клеток. Все фоторецепторы, посылающие сигналы к одной ГК, составляют ее рецепторное поле. Рецепторные поля, образованные П, гораздо многочисленнее, по сравнению с К рецепторными полями. Необходимо отметить, что каждая К, находящаяся в центральной ямке желтого пятна сетчатки, связана только с одной ГК. Другими словами, рецепторное поле нейрона включает только одну колбочку. С этим связана наилучшая разрешающая способность центрального К зрения. Другие ганглиозные клетки получают сигналы больше, чем от 1 фоторецептора. Существуют такие ГК, которые собирают сигнализацию от множества тысяч палочек, следовательно, рецепторные поля сетчатки весьма разнообразны.
фоторецепторов. Следовательно, в среднем 1 аксон ГК собирает информацию от множества рецепторных клеток. Все фоторецепторы, посылающие сигналы к одной ГК, составляют ее рецепторное поле. Рецепторные поля, образованные П, гораздо многочисленнее, по сравнению с К рецепторными полями. Необходимо отметить, что каждая К, находящаяся в центральной ямке желтого пятна сетчатки, связана только с одной ГК. Другими словами, рецепторное поле нейрона включает только одну колбочку. С этим связана наилучшая разрешающая способность центрального К зрения. Другие ганглиозные клетки получают сигналы больше, чем от 1 фоторецептора. Существуют такие ГК, которые собирают сигнализацию от множества тысяч палочек, следовательно, рецепторные поля сетчатки весьма разнообразны.
ОСНОВЫ ФОТОМЕТРИИ
Свойства ФР и всей зрительной системы человека оказали весьма большое влияние на фотометрию. Фотометрия – это раздел физической оптики, в котором рассматриваются энергетические характеристики светового излучения в процессах его испускания, распределения и взаимодействия с веществом. К области фотометрии относится только та часть электромагнитного спектра, которая воспринимается человеческим глазом. В необъятном спектре электромагнитных колебаний, только узкая полоса видимой области электромагнитных колебаний с длиной волн от 380 нм до 760 нм, воспринимается зрительной системой человеческого глаза, то есть, как принято говорить, обладает видимостью. Однако, к волнам разной длины, принадлежащим к видимой области, электромагнитная колебательная чувствительность зрительного анализатора человека неодинакова, следовательно, различаемые участки видимой области спектра, обладают разной видимостью. Кривые видности были построены в 20-е годы ХХ века по результатам определения чувствительности зрительного анализатора человека к свету разных длин волн. Кривые видности имеют вид:

Кривые видности фотопического (сплошная линия) и скотопического
(штриховая линия) зрения
Дата публикования: 2014-11-04; Прочитано: 728 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
