 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Вторичная структура ДНК
|
|
Изучая состав ДНК, Чаргафф в 1949 г. установил важные закономерности, касающиеся отдельных оснований ДНК. Они помогли раскрыть вторичную структуру ДНК. Эти закономерности называют правилами Чаргаффа:
1) сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклеотидов, т.е. А+ГУЦ+Т = 1;
2) содержание аденина равно содержанию тимина: А=Т;
3) содержание гуанина равно содержанию цитозина: Г=Ц;
4) количество аденина и цитозина равно количеству гуанина и тимина: А+Ц=Г+Т;
5) изменчивы только суммы А+Т и Г+Ц. Если А+Т>Г+Ц, то это AT - тип ДНК; если Г+Ц>А+Т, то это ГЦ-тип ДНК.
Эти правила говорят о том, что при построении ДНК должно соблюдаться довольно строгое соответствие не пуриновых и пиримидиновых оснований вообще, а конкретно тимина с аденином и цитозина с гуанином.
На основании правил Чаргаффа и рентгеноструктурных исследований Дж. Уотсон и Ф. Крик (Великобритания) предложили модель строения ДНК (1953), основные черты которой сформулированы ниже.
1. Молекула ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно и на всем протяжении связанных друг с другом водородными связями.
2. Водородные связи между цепями образуются за счет специфического комплементарного взаимодействия аденинового остатка одной цепи с тиминовым остатком другой цепи (пара А - Т) и гуанинового остатка одной цепи с цитозиновым остатком другой цепи (пара Г - Ц):
Основания А и Т, Ц и Г комплементарны друг другу в том смысле, что между ними легче возникают водородные связи, чем при других сочетаниях (например, А и Г, Ц и А и т.д.).
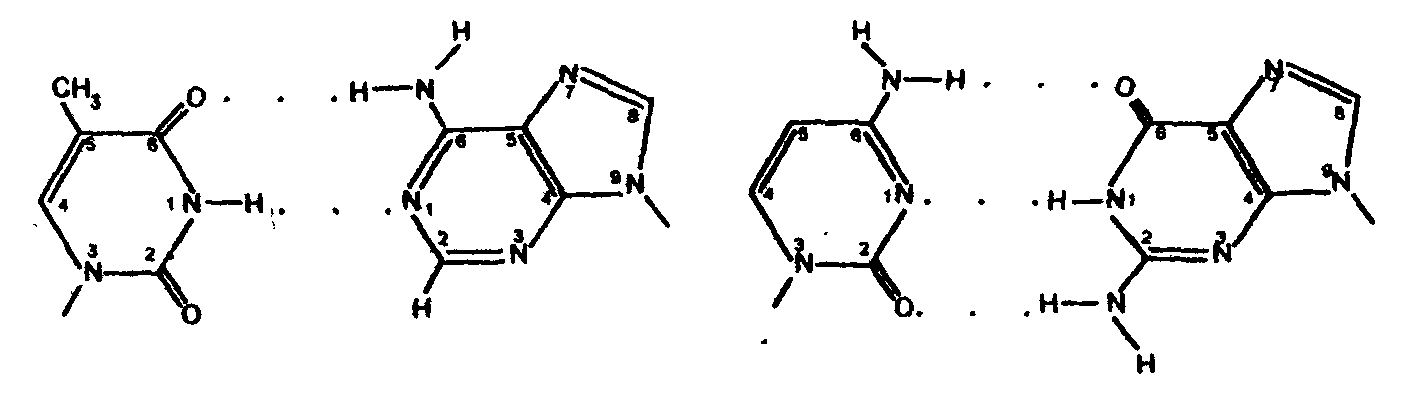
| Тимин | Аденин | Цитозин | Гуанин |
3. Первичная структура одной цепи молекулы ДНК комплементарна первичной структуре другой цепи. Это можно показать следующей схемой:

Таким образом, если известна первичная структура одной цепи, легко может быть написана, исходя из правил комплементарности, первичная структура другой цепи.
4. Обе цепи закручены в спираль, имеющую общую ось; цепи могут быть разъединены только путем раскручивания. Пуриновые и пиримидиновые основания обращены внутрь спирали; их плоскости перпендикулярны оси спирали и параллельны друг другу, образуя таким образом стопку оснований. Между основаниями в этой стопке возникают гидрофобные взаимодействия, которые стабилизируют двойную спираль даже больше, чем водородные связи между цепями. Пентозофосфатные части располагаются по периферии, образуя ковалентный остов спирали. По Уотсону и Крику: расстояние между витками (шаг спирали) равно 3,4 нм. На этом участке укладывается 10 нуклеотидных остатков, размер одного нуклеотида 0,34 нм, диаметр биспиральной молекулы равен 1,8 нм.

Рис. Схематическое изображение двойной спирали ДНК:
а - по Уотсону и Крику; б - А-форма ДНК; в - В-форма ДНК (С - остаток дезоксирибозы; Р - остаток фосфорной кислоты)
В настоящее время методами рентгеноструктурного анализа установлено существование четырех форм ДНК: А, В, С, Т. Простейшие из них А и В. У формы А наблюдается смещение пар оснований от оси молекулы к периферии, поэтому уменьшается шаг спирали до 2,8 нм, и в одном витке 11 мононуклеотидов вместо 10. В-форма приближается к модели Уотсона и Крика: шаг спирали 3,4 нм, диаметр биспирали - 1,7 нм. Между А- и В- формами ДНК осуществляются взаимопереходы, которые могут быть вызваны растворителями или белками. В этих переходах, очевидно, заключен определенный биологический смысл. Предполагается, что в А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК на матрице ДНК), а в В-форме - роль матрицы в процессе репликации (синтез ДНК на матрице родительской ДНК).
Установлено, что в клетках ряда бактерий и вирусов ДНК существует в виде одноцепочечной молекулы, вторичная структура которой представляет собой сочетание неспирализованных участков с участками спирали, образованной за счет взаимодействия комплементарных друг другу оснований.
Описанная структура ДНК позволяет объяснить молекулярный механизм функциональных биологических явлений, таких как самовоспроизведение организмов, наследственность, изменчивость. Поэтому. 1953 г., когда Ф. Крик и Дж. Уотсон разработали модель строения ДНК, принято считать годом рождения молекулярной биологии.
Дата публикования: 2015-11-01; Прочитано: 2191 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
