 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Фосфатидилинозитол
|
|
Фосфатидилинозитолы довольно широко распространены в природе. Они обнаружены у животных, растений и микроорганизмов. В животном организме найдены в мозге, печени и легких.
Плазмалогены. От рассмотренных глицеролипидов плазмалогены отличаются тем, что вместо одного остатка высшей жирной кислоты содержат остаток α,β-ненасыщенного спирта, который образует простую связь (в отличие от сложноэфирной связи, образуемой остатком жирной кислоты) с гидроксильной группой глицерина в положении С-1:
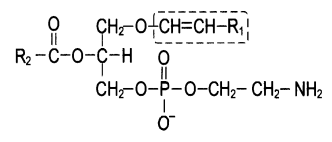
Фосфатидальхолин (плазмалоген)
Основными подклассами плазмалогенов являются фосфатидальхолины, фосфатидальэтаноламины и фосфатидальсерины. В разбавленных кислотах они гидролизуются с образованием альдегида соответствующего α,β-ненасыщенного спирта, то есть при кислотном гидролизе плазмалогенов образуются «жирные» альдегиды, называемые плазмалями, что и легло в основу термина «плазмалоген». Плазмалогены также входят в состав мембран мышц, нервных клеток, эритроцитов.
Некоторые, открытые сравнительно недавно фосфатиды, не содержат азотистого основания, место которого в молекуле в этом случае занимают глицерин и его производные:
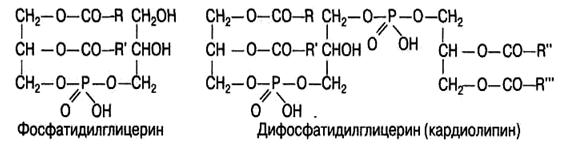
Фосфатидилглицерин является обязательной составной частью хлоропластов и в небольших количествах присутствует в бактериальных клетках и тканях животных.
Кардиолипин — одно из необходимых соединений в составе митохондриальных мембран, особенно в митохондриях сердечной мышцы.
5.1.2. Сфинголипиды
Другой группой фосфолипидов являются сфинголипиды. В их состав вместо глицерина входит высший двухатомный ненасыщенный аминоспирт сфингозин (число углеродных атомов 18, двойная связь имеет транс-конфигурацию). Гидроксильные группы расположены у первого и третьего углеродного атомов, аминогруппа находится при втором атоме углерода цепи, двойная связь - при четвертом атоме углерода:

Сфингозин
Отличие сфинголипидов от глицерофосфолипидов и в том, что жирные кислоты соединяются в них не с гидроксильными группами, а с аминогруппами аминоспирта, образуя амидные связи (-СО-NH-).Эта связь образуется между карбоксильной группой жирной кислоты и аминогруппой спирта. В сфинголипидах в значительных количествах обнаружены лигноцериновая и нервоновая кислоты.
При гидроксильной группе в положении 1 в сфинголипидах имеется остаток фосфорной кислоты, который в свою очередь этерифицирован с молекулой азотистого соединения - чаще всего с холином. Общая структура сфингомиелинов выглядит так:
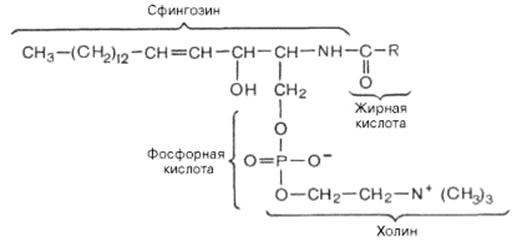
Cфингомиелин
Общий план построения молекулы сфингомиелина в определенном отношении напоминает строение глицерофосфолипидов. Молекула сфин-гомиелина содержит как бы полярную «головку», которая несет одновременно и положительный (остаток холина), и отрицательный (остаток фосфорной кислоты) заряды, и два неполярных «хвоста» (длинная алифатическая цепь сфингозина и ацильный радикал жирной кислоты).
Сфингомиелины являются самыми распространенными сфинголипидами. Они находятся, в основном, в мембранах животных и растительных клеток. Особенно богата ими нервная ткань. Собственно название сфингомиелины отражает их функцию в живых тканях, где они образуют миелиновую оболочку вокруг нервных волокон в тканях печени, почек и других органов.
В некоторых сфингомиелинах, например выделенных из мозга и селезенки, вместо сфингозина найден спирт дигидросфингозин (восстановленный сфингозин):
 СН3-(СН2)14-СН-СН-СН2-СН2ОН
СН3-(СН2)14-СН-СН-СН2-СН2ОН
ОН NH2
Дегидросфингозин
Из фосфолипидов растительного происхождения (из кукурузного зерна) выделен аминоспирт, весьма похожий на сфингозин:

Фитосфингозин
Этот же спирт найден в дрожжах и грибах, а недавно в мозге и почках человека, что указывает на возможность существования аналогичных фосфолипидов в растительных и животных объектах.
Сфингофосфолипиды не растворимы в серном эфире, что используют при их отделении от фосфатидов. Они характеризуются также трудной растворимостью в ацетоне и большей устойчивостью к действию окислителей, чем фосфатиды.
Сфинголипидам свойственны весьма сложные пространственные конфигурации, связанные с возможностью оптической изомерии (два асимметричных углеродных атома в молекуле) и цис-транс-изомерии по месту двойной связи. Этим объясняется их органная и видовая специфичность. Кроме того, установлено, что органная специфичность сфинголипидов зависит от качественного состава высших жирных кислот: так, для сфинголипидов мозга характерно присутствие нервоновой кислоты.
5.2.Гликолипиды
Вторую группу сложных липидов образуют гликолипиды (от греч. glykys-сладкий и липиды). Они характеризуются тем, что полярная моно- или олигосахаридная составляющая (глюкоза, галактоза, глюкозамин, галактозамин, их N-ацетильные производные и др.) через остаток многоатомного спирта (глицерин, сфингозин) соединяется с неполярными радикалами высших жирных кислот (пальмитиновой, стеариновой, олеиновой, лигноцериновой, нервоновой, цереброновой и др.) гликозидной исложноэфирной связями.
В зависимости от природы липидной части гликолипиды можно разделить на четыре группы:
1) гликозилдиглицериды, липидная часть которых представляет собой остаток глицерина, ацилированный в положения 1 и 2 высшими жирными кислотами;
2) гликосфинголипиды, в которых липидным фрагментом является церамид - остаток высшего аминоспирта (сфингозинового основания), N-ацилированного высшей жирной кислотой;
3) полипренилфосфатсахара, у которых липидная часть молекулы представляет собой остаток полипренола Н(СН2С(СН3)=СНСН2)nОН;
4) гликолипиды микроорганизмов, в состав которых входят остатки высших жирных кислот, ацилирующих остатки сахаров или неуглеводных компонентов молекулы.
Подавляющее большинство гликолипидов относятся к первым двум группам и являются важными компонентами мембран.
Церамиды — основа гликолипидов. Водород в гидроксильной группе церамида может быть замещён на разные углеводные фрагменты, что определяет принадлежность гликолипида к определённому классу.
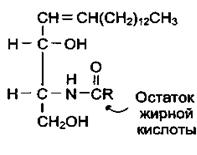
Церамид (N-ацилсфингозин)
Гликолипиды играют существенную роль в функционировании биологических мембран, содержатся преимущественно в ткани мозга, но имеются также в кровяных клетках и других тканях.
В животных системах широко распространены следующие группы гликолипидов: цереброзиды, ганглиозиды, сульфатиды. Названия «цереброзиды» и «ганглиозиды» указывают на ткани, откуда они впервые были выделены.
5.2.1. Цереброзиды
Цереброзиды (галактозилцерамиды, глюкозилцерамиды) на первый взгляд похожи на сфингомиелины, так как они не содержат ни фосфорной кислоты, ни холина. Остаток фосфорной кислоты, азотсодержащего основания замещен молекулой моносахарида - галактозы (реже глюкозы).
Галактозилцерамиды – главные сфинголипиды мозга и других нервных тканей, но в небольших количествах встречаются и во многих других тканях. В состав галактозилцерамидов входит гексоза (обычно это D-галактоза), которая связана эфирной связью с гидроксильной группой аминоспирта сфингозина. Кроме того, в составе галактозилцерамида имеется жирная кислота. Чаще всего это лигноцериновая, нервоновая или це-реброновая кислота, то есть жирные кислоты, имеющие 24 углеродных атома.
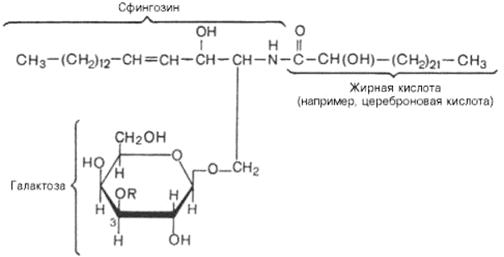
Галактозилцерамид
Глюкозилцерамиды – простые гликосфинголипиды, представлены в тканях, отличных от нервной, причем главным образом глюкозил-церамидами. В небольших количествах они имеются в ткани мозга. В отличие от галактозилцерамидов у них вместо остатка галактозы имеется остаток глюкозы.
5.2.2. Сульфатиды
Сульфатиды отличаются от цереброзидов тем, что третий углеродный атом гексозы этерифицирован серной кислотой. Сульфатиды обладают свойствами кислот и поэтому называются кислыми сфинголипидами. При физиологических значениях рН сульфатированный углеводный остаток имеет отрицательный заряд. Около 25% цереброзидов мозга представляют собой сульфатированные производные.
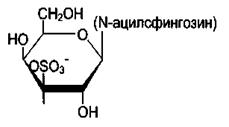
Сульфатид
Сульфатиды в значительных количествах находят в белом веществе мозга, но при этом их содержание ниже, чем цереброзидов.
5.2.3. Ганглиозиды
Особую группу гликолипидов составляют ганглиозиды – сложные, богатые углеводами липиды с молекулярной массой около 250000. В структурном отношении ганглиозиды сходны с цереброзидами. Но они дополнительно содержат одну или несколько молекул сиаловой кислоты. В тканях человека доминирующей сиаловой кислотой является нейраминовая. Она представляет собой углевод, состоящий из 9 атомов углерода. Кроме того, вместо остатка глюкозы они чаще содержат сложный олигосахарид.
Одним из простейших ганглиозидов является гематозид, выделенный из стромы эритроцитов. Он содержит церамид (ацилсфингозин), одну молекулу глюкозы, одну молекулу галактозы и одну молекулу N-ацетилнейраминовой кислоты:
Продуктами гидролиза ганглиозидов, выделенных из мозга быка, являются жирные кислоты, сфингозин, сахара (Д-глюкоза, Д-галактоза) и производные аминосахаров (N-ацетилглюкозамин, N-ацетилнейраминовую кислоту).
Ганглиозиды - важные компоненты специфических рецепторных участков, расположенных на поверхности клеточных мембран. Они находятся в тех участках нервных окончаний, где происходит связывание молекул нейромедиатора в процессе химической передачи импульса от одной нервной клетки к другой. Таким образом, ганглиозиды обеспечивают нормальное, функционирование нервной системы.
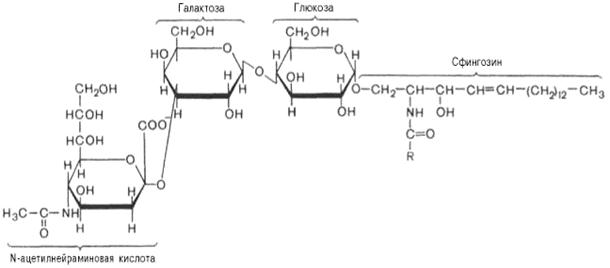
Гематозид (ганглиозид)
Большинство из рассмотренных групп сложных липидов выполняют в клетках, прежде всего, структурную функцию, являясь составными компонентами мембран.
Несмотря на большое разнообразие структур, гликолипиды обладают некоторыми общими свойствами. Они являются ПАВ: благодаря присутствию длинных алифатических цепочек, они могут растворятся в некоторых орг. растворителях (например, в бутаноле, смеси хлороформа и метанола), а наличие углеводной части придает им растворимость в воде, возрастающую с увеличением длины углеводной цепи.
6. Строение и функции биологических мембран
Биологические мембраны - общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.). Они содержат в своем составе липиды, белки,гетерогенные молекулы (гликопротеины,гликолипиды)и в зависимости от выполняемой функции многочисленные минорные компоненты: коферменты, нуклеиновые кислоты, антиоксиданты, каротиноиды, неорганические ионы и т. п.
Согласованное функционирование мембранных систем – рецепторов, ферментов, транспортных механизмов - помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям биологических мембран можно отнести:
· отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
· контроль и регулирование транспорта огромного многообразия веществ через мембраны;
· участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
· преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
Молекулярная организация плазматической (клеточной) мембраны у всех клеток примерно одинакова: она состоит из двух слоев липидных молекул с множеством включенных в нее специфических белков. Одни мембранные белки обладают ферментативной активностью, тогда как другие связывают питательные вещества из окружающей среды и обеспечивают их перенос в клетку через мембраны. Мембранные белки различают по характеру связи с мембранными структурами. Одни белки, называемые внешними или периферическими, непрочно связаны с поверхностью мембраны, другие, называемые внутренними или интегральными, погружены внутрь мембраны. Периферические белки легко экстрагируются, тогда как интегральные белки могут быть выделены только при помощи детергенов или органических растворителей. На рис. 4 представлена структура плазматической мембраны.
Внешние, или плазматические, мембраны многих клеток, а также мембраны внутриклеточных органелл, например, митохондрий, хлоропластов удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембран от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах клеток мозга наоборот, около 80% липидов и только 20% белков.
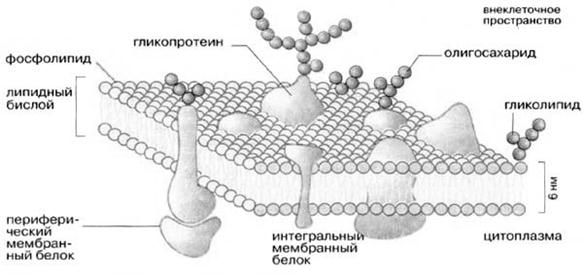
Рис. 4. Структура плазматической мембраны
Липидная часть мембран представляет собой смесь различного рода полярных липидов. Полярные липиды, к числу которых относятся фосфоглицеролипиды, сфинголипиды, гликолипиды не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях.
Все полярные липиды в мембранах постоянно обновляются в процессе метаболизма, при нормальных условиях в клетке устанавливается динамическое стационарное состояние, при котором скорость синтеза липидов равна скорости их распада.
В мембранах животных клеток присутствуют в основном фосфоглицеролипиды и в меньшей степени сфинголипиды; триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержит значительные количества холестерола и его эфиров (рис.5).
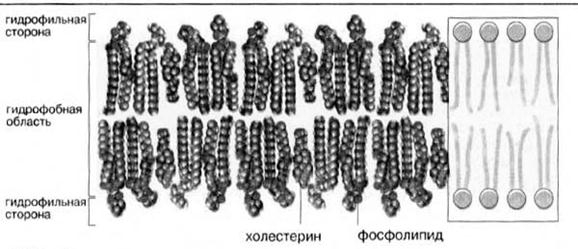
Рис.5. Мембранные липиды
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 году С. Синджером и Дж. Николсоном.
Согласно ей белки можно уподобить айсбергам, плавающим в липидном море. Как уже указывалось выше, существуют 2 типа мембранных белков: интегральные и периферические. Интегральные белки пронизывают мембрану насквозь, они являются амфипатическими молекулами. Периферические белки не пронизывают мембрану и связаны с ней менее прочно. Основной непрерывной частью мембраны, то есть ее матриксом, служит полярный липидный бислой. При обычной для клетки температуре матрикс находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных липидов.
Жидкостно-мозаичная модель предполагает также, что на поверхности расположенных в мембране интегральных белков имеются R-группы аминокислотных остатков (в основном гидрофобные группы, за счет которых белки как бы «растворяются» в центральной гидрофобной части бислоя). В то же время, на поверхности периферических, или внешних белков, имеются в основном гидрофильные R-группы, которые притягиваются к гидрофильным заряженным полярным головкам липидов за счет электростатических сил. Интегральные белки, а к ним относятся ферменты и транспортные белки, обладают активностью только в том случае, если находятся внутри гидрофобной части бислоя, где они приобретают необходимую для проявления активности пространственную конфигурацию (рис.6). Следует еще раз подчеркнуть, что ни между молекулами в бислое, ни между белками и липидами бислоя не образуется ковалентных связей.


Рис.6. Мембранные белки
Мембранные белки могут свободно перемещаться в латериальной плоскости. Периферические белки буквально плавают на поверхности бислойного «моря», а интегральные белки, подобно айсбергам, почти полностью погружены в углеводородный слой.
В большинстве своем мембраны ассиметричны, то есть имеют неравноценные стороны. Эта ассиметричность проявляется в следующем:
· во-первых, в том, что внутренняя и внешняя стороны плазматических мембран бактериальных и животных клеток различаются по составу полярных липидов. Так, например, внутренний липидный слой мембран эритроцитов человека содержит в основном фосфатидилэтаноламин и фосфатидилсерин, а внешний – фосфатидилхолин и сфингомиелин.
· во-вторых, некоторые транспортные системы в мембранах действуют только в одном направлении. Например, в мембранах эритроцитов имеется транспортная система («насос»), перекачивающая ионы Nа+ из клетки в окружающую среду, а ионы К+ - внутрь клетки за счет энергии, освобождающейся при гидролизе АТФ.
· в-третьих, на внешней поверхности плазматических мембран содержится очень большое число олигосахаридных группировок, представляющих собой головки гликолипидов и олигосахаридные боковые цепи гликопротеинов, тогда как на внутренней поверхности плазматической мембраны олигосахаридных группировок практически нет.
Ассиметричность биологических мембрам сохраняется за счет того, что перенос индивидуальных молекул фосфолипидов с одной стороны липидного бислоя на другую очень затруднен по энергетическим соображениям. Полярная молекула липида способна свободно перемещаться на своей стороне бислоя, но ограничена в возможности перескочить на другую сторону.
Подвижность липидов зависит от относительного содержания и типа присутствующих ненасыщенных жирных кислот. Углеводородная природа жирнокислотных цепей сообщает мембране свойства текучести, подвижности. В присутствии цис-ненасыщенных жирных кислот силы сцепления между цепями слабее, чем в случае одних насыщенных жирных кислот, и липиды сохраняют высокую подвижность и при низкой температуре.
На внешней стороне мембран имеются специфические распознающие участки, функция которых состоит в распознавании определенных молекулярных сигналов. Например, именно посредством мембраны некоторые бактерии воспринимают незначительные изменения концентрации питательного вещества, что стимулирует их движение к источнику пищи; это явление носит название хемотаксиса.
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов:
· плазматическая мембрана (наружная клеточная мембрана, плазмалемма),
· ядерная мембрана,
· эндоплазматический ретикулум,
· мембраны аппарата Гольджи, митохондрий, хлорпластов, миелиновых оболочек,
· возбудимые мембраны.
У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются впячиванием внуть наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Мембрана эритроцитов состоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная – с липидами. В мембране липиды расположены асимметрично в отличие от симметричного расположения в мицеллах. Например, кефалин находится преимущественно во внутреннем слое липидов. Такая асимметрия поддерживается, по-видимому, за счет поперечного перемещения фосфолипидов в мембране, осуществляемого с помощью мембранных белков и за счет энергии метаболизма. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружном слое – фосфатидилхолин. Мембрана эритроцитов содержит интегральный гликопротеин гликофорин, состоящий из 131 аминокислотного остатка и пронизывающий мембрану, и так называемый белок полосы 3, состоящий из 900 аминокислотных остатков. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов. В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин.
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них – фосфолипидов). Белки этих мембран важны для фиксации лежащих друг над другом мембранных солев.
Мембраны хлоропластов. Хлоропласты покрыты двухслойной мембраной. Наружная мембрана имеет некоторое сходство с таковой у митохондрий. Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система – ламеллы. Ламеллы образуют или уплощенные пузырьки – тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте в гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, активного транспорта; ферменты, участвующие в образовании компонентов мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3:1. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Мембрана вирусов. Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (микровирусы) в мембраны входит 70-80% всех белков, остальные белки содержатся в нуклеокапсиде.
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, что придает поверхности мембран биологическую специфичность.
7. КЛЮЧЕВЫЕ ТЕРМИНЫ И ПОНЯТИЯ К ГЛАВЕ I
Арахидоновая кислота (С19Н31СООН) - одноосновная, ненасыщенная кислота с четырьмя двойными связями; исходное соединение для синтеза большой группы биологически активных веществ - эйкозаноидов (простагландинов, тромбоксанов, лейкотриенов).
Биологические мембраны - общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.).
Воски — простые липиды, являющиеся сложными эфирами высших спиртов и высших монокарбоновых кислот. Природные В. содержат также свободные высшие жирные кислоты, высшие спирты, триглицериды и углеводороды. В. - твердые вещества разнообразной окраски, обладают водоотталкивающими свойствами, приобретают пластичность при незначительном нагревании (температура плавления от 30 до 90 °С).
Высшие жирные кислоты ( В.ж. к.) - органические соединения, представляющие собой монокарбоновые кислоты алифатического ряда с числом углеродных атомов в молекуле, равным 14 и более. Основной структурный компонент многих липидов (жиров, восков, стероидов, глико-, фосфо- и диольных липидов). В. ж. к. могут быть насыщенными (пальмитиновая, стеариновая и др. кислоты) и ненасыщенными, содержащими одну (олеиновая кислота) и более двойных связей (линолевая, линоленовая, арахидоновая кислоты), тройную связь (тарариновая кислота), а также оксигруппы в углеводородном радикале. В составе природных липидов преобладают В. ж. к. нормального строения с четным числом углеродных атомов в молекуле.
Ганглиозиды - гликолипиды, содержащие в своем составе несколько углеводных остатков (среди которых присутствует N-ацетилнейраминовая кислота), связанных с церамидом. Содержатся в ганглиозных клетках нервной ткани, в плазматических мембранах многих клеток; принимают участие в межклеточных контактах.
Гликолипиды - сложные липиды, содержащие в своем составе углеводные компоненты. Широко распространены в природе, находятся в основном в мембранах клеток нервной ткани; они регулируют рост клеток, играют важную роль в явлениях клеточной адгезии, участвуют в процессах рецепции гормонов, являются иммуномодуляторами и вторичными посредниками.
Глицерин (глицерол, пропантриол-1,2,3) - трехатомный спирт, структурный компонент жиров, глицерофосфолипидов.
Глицерофосфолипиды, фосфатиды - сложные эфиры глицерина, высших жирных кислот, фосфорной кислоты и азотистого основания. В зависимости от характера азотистого основания различают фосфатидилхолины, фосфатидилколамины, фосфатидилсерины и фосфатидилтреонины.
Жиры, триглицериды- сложные эфиры трехатомного спирта глицерина и одноосновных высших жирных кислот чаще всего с четным числом атомов углерода. Ж. относятся к нейтральным липидам. Простые Ж. - сложные эфиры глицерина и одной из высших жирных кислот, смешанные Ж. -сложные эфиры глицерина и остатков разных высших жирных кислот. Ж. - энергетическое депо, субстраты для окисления и обеспечения организма энергией; располагаются преимущественно в подкожной жировой ткани и выполняют функции теплоизоляционной и механической защиты.
Инозитфосфолипиды, фосфоинозитиды- фосфолипиды, сложные эфиры шестиатомного циклогексанового спирта миоинозитола и фосфатидных кислот. Могут содержать в своем составе два или три остатка фосфорной кислоты, моносахариды, амины, аминокислоты, жирные кислоты - все они присоединяются к остатку инозитола. И. содержатся в растительных и животных (особенно нервных) тканях, являясь постоянными компонентами липидного бислоя биологических мембран.
Линолевая кислота (С17Н31СООН) - одноосновная, незаменимая, ненасыщенная, с двумя двойными связями (диеновая) карбоновая кислота; предшественник арахидоновой кислоты.
Линоленовая кислота (С17Н29СООН) - одноосновная, незаменимая, ненасыщенная, с тремя двойными связями (триеновая) карбоновая кислота. Л. к. в виде триглицеридовсодержится в льняном, конопляном и др. маслах.
Липиды - природные органические вещества, нерастворимые в воде, но растворимые в жирорастворителях, являются производными высших жирных кислот.Л. бывают простые и сложные. Простые Л. представлены двухкомпонентными веществами - сложными эфирами высших жирных кислот с глицерином (жиры), высшими (воски) или полициклическими (стериды) спиртами. Сложные Л. - многокомпонентные молекулы, к ним принадлежат фосфолипиды, гликолипиды, диольныелипиды, орнитинолипиды.
Олеиновая кислота (С17Н33СООН) - непредельная жирная кислота; содержится в виде глицеридов во всех растительных маслах и животных жирах.
Пальмитиновая кислота (С15Н31СООН) - предельная одноосновная карбоновая кислота; входит в состав ацилглицеридов всех животных жиров и растительных масел, в состав некоторых восков (например, спермацета, пчелиного воска).
Стеариновая кислота (С17Н35СООН) - насыщенная высшая жирная кислота. Входит в состав всех животных жиров и растительных масел, в состав глико- и фосфолипидов различного происхождения (обычно вместе с пальмитиновой кислотой).
Стериды - группа простых липидов, по химической природе сложные эфиры циклических спиртов(стеролов) и высших жирных кислот, из которых в составе С. обнаружены пальмитиновая, стеариновая, олеиновая и другие кислоты. У животных основным С. является холестерид - важный компонент липопротеинов плазмы, а также наружной клеточной мембраны; входит в состав нервной ткани. В мембранах растительных клеток содержатся стигма- и ситостериды.
Стероиды - класс органических полициклических соединений, широко распространенных в живой природе. В основе С. лежит остов циклопентанопергидрофенантрена. Общий биогенетический предшественник С. -сквален.Для С. характерно присутствие гидроксильной или кетогруппы в положении С3 молекулы, то есть они являются окисленной формой стеролов.
Стеролы - циклические спирты, химическую основу которых составляет циклопентанопергидрофенантрен (стеран), несущий боковую связь углеродных атомов и две метильные группы (при 10- и 13-м углеродных атомов цикла) - холестан.
Сфингозин (транс-1,3-диокси-2-амино-октадецен-4) - высший алифатический ненасыщенный аминоспирт, основной структурный компонент сфингофосфолипидов и гликолипидов.
Сфингофосфолипиды - фосфолипиды, содержащие двухатомный аминоспирт сфингозин, ацилированный по аминогруппе остатком высшей жирной кислоты (чаще лигноцериновой, нервоновой - это кислоты с длинной цепью - 24 атома углерода), по первичной гидроксильной группе - остатком фосфорной кислоты, которая, в свою очередь, соединена с холином, либо с этаноламином, либо с глицерином. С. - основные компоненты миелина и мембран клеток мозга и нервной ткани.
Фосфатидилколамины, кефалины, фосфатидилэтаноламины - фосфолипиды, сложные эфиры фосфатидных кислот и аминоспирта этаноламина (коламина). Различаются Ф. входящими в их состав высшими жирными кислотами. Являются основными липидными компонентами биологических мембран.
Фосфатидилсерины - фосфолипиды, сложные эфиры фосфатидных кислот и аминокислоты серина. Являются минорными компоненты липидного бислоя биологических мембран, специфическими активаторами Na+/К+-АТФ-азы и Са2+-фосфолипидзависимой протеинкиназы С, осуществляющей фосфори- лирование ряда ферментов.
Фосфатидилхолины, лецитины - фосфолипиды, сложные эфиры фосфатидных кислот и аминоспирта холина. Наиболее распространенные в природе фосфолипиды содержатся в тканях животных (головной мозг, эритроциты, яичный желток), растений (бобы сои, семена подсолнечника), дрожжах. Разнообразие Ф. определяется строением входящих в их состав высших жирных кислот. Ф. - главный компонент липидного слоя биологических мембран.
Фосфатидные кислоты - природные соединения из группы фосфолипидов; простейшие представители глицерофосфолипидов. Можно рассматривать как производные α-глицерофосфата, две гидроксильные группы которого замещены остатками высших жирных кислот. Присутствуют в незначительных количествах в животных и растительных тканях, являются важными промежуточными соединениями в биосинтезе триглицеридов и глицерофосфолипидов.
Фосфолипиды - сложные эфиры многоатомных спиртов с высшими жирными кислотами, содержащие остатки фосфорной кислоты и связанные с ней добавочные группировки (азотистые основания, аминокислоты, глицерин, миоинозит, сфингозин). Из многоатомных спиртов в составе различных Ф. найдено три: глицерин (глицерофосфолипиды), миоинозит (инозит- фосфолипиды), сфингозин (сфингофосфолипиды). Разнообразие Ф. определяется природой не только спиртовых компонентов, но и остатков высших жирных кислот. Ф. широко распространены в живой природе, легко образуют комплексы с белками и в виде фосфолипопротеинов участвуют в формировании клеточной оболочки и внутриклеточных мембран.
Холестерол, холестерин— полициклический одноатомный непредельный вторичный спирт, важнейший зоостерол (С27). В основе X. лежит остов стерана. X. - предшественник стероидных гормонов, желчных кислот и витамина D3 (холекальциферола). Он содержится во всех тканях животного организма, главным образом в нервной ткани, в печени, в надпочечниках и в эритроцитах. X. - структурный компонент всех клеточных мембран и липопротеинов плазмы крови. Концентрация X. в крови взрослых людей составляет 200 мг %, или 5,2 ± 1,2 ммоль/л, и, как правило, увеличивается с возрастом.
Холин -триметильное производное этаноламина, входит в состав глицерофосфолипида фосфатидилхолина, относится к витаминам.
Дата публикования: 2014-09-25; Прочитано: 7335 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
