 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Глава 3. Стандартный обмен и критерий упорядоченности у позвоночных животных
|
|
Класс Amphioxi (Ланцетники)
Ланцетники, по мнению зоологов, являются наиболее древними животными в типе позвоночных. Исходя из этого, было бы желательным иметь как можно больше доброкачественных данных о стандартном обмене этих животных. К сожалению, это не так. Нам известны только четыре работы, в которых приводятся данные о стандартном обмене ланцетника Branchiostoma lanceolatum (Ивлев, 1961; Courtney, Newell, 1965; Ивлева, Попен кина, 1968, Ивлева, 1981). В трех последних работах приводятся сухая, но не сырая масса тела ланцетников, поэтому для расчета стандартного обмена мы использовали данные о содержании воды в теле ланцетников равной 78.5% (Виноградов, 1937). Расчет по данным указанных авторов показывает, что для Branchiostoma lanceolatum α=0.352, Cr =5.37 (Courtney, Newell, 1965), a=0.258, Cr=3.94 (Ивлева, Попе нченко, 1969) и а=0.301, Сr=6.84 (Ивлева, 1981). С другой стороны, расчет по данным Ивлева (1961) показывает, что для этого вида ланцетника а=1.375, Сr=20.98. В целом без учета данных Ивлева имеем предварительные величины
Класс Amphioxi a =0.304; Cr =5.38 (118)
Очевидно, что предстоит еще большая работа для того, чтобы получить правильные величины а и Сr для класса Amphioxi.
Рыбы
В состав этой группы в соответствии с принятой в настоящее время классификацией рыб (Решетников и др., 1989), мы включили четыре класса: Cephalaspidomorphi (Миноги), Myxini (Миксинообразные), Chondrichthyes (Хрящевые рыбы) и Osteichthyes (Костные рыбы).
Основная трудность изучения стандартного обмена у рыб, помимо всего прочего, связана с тем, что разные рыбы обладают разной степенью подвижности и при измерении скорости потребления кислорода рыбами исследователи сталкиваются с разной величиной вклада активного обмена в получаемую величину стандартного обмена. Избежать этой неточности очень трудно, поэтому во многих случаях мы имеем дело не со стандартным, а активным обменом, что приводит к завышению величины коэффициента а. Чтобы снизить величину ошибки при определении этого коэффициента можно было бы для активно плавающих рыб ввести поправку на активный обмен. Для этого можно было бы определять у рыб не стандартный, а "основной" обмен - обмен у неподвижных или наркотизированных рыб (Винберг, 1956). Оказалось, что чем больше подвижность рыб во время опыта, тем больше отличие стандартного обмена у нормальных и наркотизированных рыб: у разных видов рыб в зависимости от их подвижности "основной" обмен составляет от 40 до 100% от стандартного (Белокопытин, 1973). Следовательно, для внесения поправки необходимо иметь данные о дыхании неподвижных или наркотизированных рыб. Таких данных в литературе слишком мало, поэтому мы не можем учитывать это обстоятельство в своих расчетах, что, конечно, делает весьма условным приводимую нами классификацию рыб по стандартному обмену.
Следующая неточность связана с отсутствием достоверных данных о содержании воды в теле рыб. В сводке Виноградова (1937) приводятся многочисленные измерения содержания воды в теле рыб, но речь идет только о мягких частях тела. Поэтому при вычислении критерия упорядоченности у рыб,
как и у других позвоночных животных, мы приняли, содержание воды в теле равным 70%.
Данных о стандартном обмене рыб накопилось в литературе довольно много. Они обобщены в монографии Винберга (1956), где приведены данные о массе тела и скорости потребления кислорода 106 видов рыб (n=350) и в статье Озернюка и Булгаковой (1997) - 178 видов рыб (n=232). К сожалению монография Г.Г.Винберга написана слишком давно и в ней, естественно, не учтены работы последних 40 лет. В статье Озернюка и Булгаковой отражены более новые работы, но в ней не приведены данные о массе тела рыб, что не позволило нам пересчитать приводимые в этой работе величины коэффициента а с помощью принятого в данной монографии коэффициента k=0.75 (авторы приняли для своих расчетов k=0.87). Нам пришлось поэтому самим собрать данные о стандартном обмене рыб. Всего нами использованы данные о стандартном обмене 236 видов рыб (n=475).
При расположении и наименовании разных таксонов рыб мы использовали работы Расса (1971), а также Решетникова и др.(1989).
Классы Myxini (Миксинообразные) и Cephalaspidomorphi (Миноги).
Согласно расчетам Винберга (1956) аллометрическая зависимость скорости потребления кислорода от массы тела при 20°С у круглоротых (N=4), так же, как и костных рыб, имеет вид
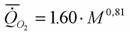
(здесь, как и везде мы выражаем коэффициент а в мВт).
Озернюк и Булгакова (1997) на основании данных о стандартном обмене 10 видов круглоротых получили несколько иные коэффициенты в аллометрической зависимости:

Значения аллометрических коэффициентов, приводимые Винбергом для круглоротых, по-видимому, имеют большую статистическую погрешность, так как при их вычислении автор использовал данные о стандартном обмене всего 4 видов миног (n=6). Озернюк и Булгакова также нашли в литературе
недостаточное число данных (3 вида миксин и 7 видов миног). Кроме того, при расчете коэффициентов в аллометрической зависимости Озернюк и Булгакова использовали не только данные о стандартном обмене взрослых миног, но и их личинок, что нельзя делать, так как пескорейки имеют значительно более низкий обмен, чем взрослые формы [табл. 34(1)].
Найденные нами материалы по дыханию круглоротых представлены в табл.34 (1). Расчеты по этим данным дают следующие значения аллометрических коэффициентов:
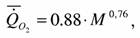
где k=0.764+0.113 достоверно не отличается от стандартной величины k=0.75. Поэтому, при расчетах коэффициента а мы использовали k=0.75.
При вычислении критерия упорядоченности мы считали, что содержание воды в теле круглоротых, как и у других позвоночных животных, близко 70%. С учетом сказанного имеем для круглоротых:
Класс Myxini a=0.237±0.094; Сr=4.23±1.68 (119)
Класс Cephalaspidomorphi a =1.025±0.128; Cr=15.27±1.91 (120)
взрослые формы a=1.440±0.095; Cr=21,45±1.42
Таблица 34(1)
Масса тела М(г), скорость потребления кислорода Q (мл О2/ч) при температуре t°C, сопоставимый стандартный обмен (коэффициент а, мВт) при 20 С и критерий упорядоченности (Cr) y рыб (классы: Myxini - Миксинообразные и Cephalaspidomorphi -Миноги).
| Классы и виды | M | Q | t | Автор | а | Cr |
| Myxini - Миксинообразные | 0.237 | 4.23 | ||||
| Eptatretus deani | 0.20 | 3.5 | (1) | 0.143 | 3.53 | |
| Eptatretus stoutii | 61.83 | 0.56 | (2) | 0.331 | 4.93 | |
| Cephalaspidomprphi Миноги взрослые формы | 1.025±0.128 1.440±0.095 | 15.27±1.91 21.45±1.42 | ||||
| Entosphenus tridentatus | 13.84 | (3) | 1.325 | 19.74 |
| Ichthyomyzon fossor | 3.78 | 0.40 | (4) | 0.992 | 14.78 | |
| Ichthyomyzon hubbsi | 0.156 | 15.5 | (5) | 0.455 | 6.78 | |
| Lampetra planeri | 5.2 | 0.79 | (6) | 1.286 | 19.17 | |
| 1.34 | 0.067 | (7) | 0.461 | 6.87 | ||
| 2.08 | 0.126 | (8) | 0.951 | 14.17 | ||
| Lampetra fluviatilis | 3.52 | 16.3 | (9) | 1.800 | 26.82 | |
| 1.27 | 0.065 | (8) | 0.710 | 10.57 | ||
| 2.50 | 12.7 | (10) | 1.198 | 17.85 | ||
| Lampetra japonica | 0.105 | (11) | 0.410 | 6.10 | ||
| Petromyzon marinus | 4.88 | 0.63 | (4) | 1.273 | 18.97 | |
| 75.6 | 6.56 | (11) | 1.435 | 21.39 |
(1) Smith,Hessler, 1974; (2) Munz,Morris, 1965; (3) Johansen et al., 1973; (4) Leach, 1946; (5) Hill,Potter, 1970; (6) Ивлев, 1954; (7) Potter,Rogers, 1972; (8) Lewis,Potter, 1977; Щербаков, 1937; (10) Claridge,Potter, 1975; (11) Beamish, 1973.
Класс Chondrichthyes (Хрящевые рыбы)
В обзоре Озернюка и Булгаковой (1997) использованы данные о стандартном обмене 9 видов акуловых рыб (п=9). Принималось, что аллометрическая зависимость дыхания от массы тела у акул такая же, как и у остальных рыб, т.е. k=0.87. Исходя из этого, авторы получили для этой группы рыб а =0.215 мл О2/г•час, т.е. в нашем обозначении а =1.206 мВт.
Расчеты по собранным нами данным дают следующую аллометрическую зависимость для хрящевых рыб:
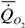 = 1.63 · М 0.76,
= 1.63 · М 0.76,
где k=0.763+0.054 достоверно не отличается от стандартной величины k= 0.75. Поэтому, при расчетах коэффициента а мы использовали k=0.75.
В сводке Виноградова (1937) приводятся данные о содержании воды в мягких тканях хрящевых рыб. Оно равно в среднем 78.5% (N=13, n=19). Так как не совсем ясно, каково содержание воды в теле акул, включая скелет, то при расчете критерия упорядоченности мы приняли, что содержание воды в теле хрящевых рыб равно 70%. Собранные нами литературные данные о стандартном
обмене хрящевых рыб и рассчитанные величины коэффициента а (принимая, что k=0.75) и критерия упорядоченности Сr показаны в табл.34(2).
Мы использовали данные о стандартном обмене 9 видов акуловых рыб (n=16) [табл. 34(2)]. При расчете критерия упорядоченности было учтено, что многие хрящевые рыбы живут в субтропической или тропической зоне океанов. Имеем:
Класс Chondrichthyes a =1.766; Сr=27.40 (121)
Таблица 34(2)
Масса тела М(г), скорость потребления кислорода Q (мл О2/ч) при температуре t°C, сопоставимый стандартный обмен (коэффициент а, мВт) при 20°С и критерий упорядоченности (Сr) у рыб (класс Chondrichthyes - Хрящевые рыбы).
| Таксон | M | Q | t | Автор | a | Cr |
| Класс Chondrichthyes | 1.766±0.099 | 27.40±1.54 | ||||
| Подкласс Elasmobranchii | 1.766±0.099 | 27.40±1.54 | ||||
| Отряд Carcharhiniformes | 1.807±0.091 | 26.54±1.34 | ||||
| Scyliorhinus cassicula | 9.62 | (1) | 1.510 | 22.50 | ||
| Scyliorhinus canicula | 8.94 | (1) | 1.957 | 29.16 | ||
| 23.7 | (2) | 1.727 | 25.80 | |||
| 8.785 | 12.2 | (3) | 2.009 | 29.93 | ||
| Scyliorhinus stellaris | 59.95 | (1) | 1.949 | 28.80 | ||
| 107.64 | (4) | 2.343 | 34.91 | |||
| 39. 19 | 18.5 | (5) | 1.886 | 28.10 | ||
| 83.71 | (6) | 2.064 | 30.76 | |||
| Scyllium cassicula | 132.6 | 4.96 | (5) | 1.311 | 19.24 | |
| Scyllium catulus | 16.78 | (5) | 1.497 | 22.30 | ||
| 21.5 | 1.776 | (5) | 1.651 | 24.73 | ||
| Отряд Orectolobiformes | 1.172 | 28.99 | ||||
| Hemiscyllium plagiosum | (7) | 1.172 | 28.99 | |||
| Отряд Squaliformes | 2.195 | 32.71 | ||||
| Squalus suckleyi | 14.91 | (8) | 2.218 | 33.05 | ||
| (1) | 2.172 | 32.36 | ||||
| Отряд Rajiformes | 1.842 | 27.68 | ||||
| Raja torpedo | 17.14 | 14.5 | (5) | 1.842 | 27.68 | |
| Отряд Torpediniformes | 0.950 | 23.49 | ||||
| Torpedo marmorata | 29.3 | (1) | 0.950 | 23.49 |
(1) Hughes, 1978; (2) Butler,Taylor, 1975; (3) Hughes,Umezawa, 1968; (4) Piiper et al., 1970; (5) Винберг, 1956; (6) Baumgarten-Schumann,Piiper, 1968; (7) Chan,Wong, 1977; (8) Gerald, Cech, 1970.
Класс Osteichthyes (Костные рыбы)
Озернюк и Булгакова (1997) приводят данные о дыхании 159 видов (n=210) костных рыб. Аллометрическая зависимость стандартного обмена от массы тела (этих и акуловых рыб) по расчетам этих авторов имеет вид:
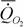 = 0.98 М 0.87.
= 0.98 М 0.87.
Мы собрали данные о стандартном обмене 228 видов рыб (n=446) и получили аллометрическую зависимость вида
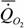 = 1.26 М 0.84,
= 1.26 М 0.84,
где k=0.844±0.080. Другими словами, коэффициент k достоверно не отличается от k=0.75.
Таблица 34(3)
Сопоставимый стандартный обмен (коэффициент а, мВт) при 20°С и критерий упорядоченности (Сr) у рыб (класс Osteichthyes - Костные рыбы). N - число изученных видов, n - число измерений.
| Таксон | а | Сr | N | n |
| Подкласс Sarcopterygii | 0.570±0.163 | 14.10±4.03 | ||
| Надотряд Dipnoi (Двоякодышащие) | 0.570±0.163 | 14.10±4.03 | ||
| Отряд Ceratodontiformes | 0.327 | 8.10 | ||
| Отряд Lepidosireniformes | 0.732 | 18.10 | ||
| Подкласс Actinopterygii | 1.634±0.231 | 24.21±3.42 | ||
| Надотряд Ganoidomorpha | 1.634±0.231 | 24.21±3.42 | ||
| Отряд Polypteriformes | 0.653 | 16.15 | ||
| Отряд Lepisosteiformes | 0.984 | 19.49 | ||
| Отряд Acipenseriformes | 1.974±0.211 | 29.42±3.14 | ||
| Подкласс Teleostei | 1.816±0.047 | 30.46±0.79 | ||
| Надотряд Batrachomorpha | 0.301 | 7.44 | ||
| Надотряд Anguillomorpha | 1.208±0.148 | 24.98±3.06 | ||
| Отряд Anguilliformes | 1.208±0.148 | 24.98±3.06 | ||
| Сем. Morhguidae | 0.306 | 7.57 | ||
| Сем. Anguillidae | 1.107±0.054 | 21.02±1.03 | ||
| Сем. Congridae | 1.715 | 42.42 | ||
| Сем. Muraenidae | 2.610 | 64.55 |
| Надотряд Cyprinomorpha | 1.639±0.067 | 25.34±1.04 | ||
| Отряд Cypriniformes | 1.774±0.071 | 26.85±1.07 | ||
| Подотряд Gymnotoidei | 0.537 | 13.28 | ||
| Сем. Gymnotidae | 0.347 | 8.59 | ||
| Сем. Electrophoridae | 0.726 | 17.96 | ||
| Подотряд Cyprinoidei | 1.808±0.071 | 27.10±1.06 | ||
| Сем. Cobitidae | 1.283±0.191 | 19.12±2.85 | ||
| Сем. Catostomidae | 1.413 | 21.05 | ||
| Сем. Cyprinidae | 1.860±0.076 | 27.89±1.14 | ||
| Отряд Siluriformes | 0.877±0.053 | 16.80±1.02 | ||
| Сем. Heteropneustidae | 0.536 | 13.26 | ||
| Сем. Ictaluridae | 0.950±0.047 | 14.16±0.70 | ||
| Сем. Loricariidae | 0.638 | 15.79 | ||
| Сем. Plotosidae | 0.651 | 16.11 | ||
| Сем. Bagridae | 0.796 | 19.68 | ||
| Сем. Saccobranchidae | 0.804 | 19.88 | ||
| Сем. Callichthyidae | 1.023 | 25.32 | ||
| Сем. Clariidae | 1.094 | 27.66 | ||
| Надотряд Atherinomorpha | 1.492±0.198 | 32.58±4.32 | ||
| Отряд Cyprinodontiformes | 1.492±0.198 | 32.58±4.32 | ||
| Сем. Poeciliidae | 1.317±0.326 | 28.00±6.93 | ||
| Сем. Cyprinodontidae | 1.461±0.119 | 36.14±2.94 | ||
| Сем. Oryziatidae | 2.903 | 43.26 | ||
| Надотряд Percomorpha | 1.837±0.080 | 32.59±1.42 | ||
| Отряд Scorpaeniformes | 1.652±0.181 | 29.27±3.21 | ||
| Подотряд Cottoidei | 1.529±0.211 | 26.51±3.66 | ||
| Сем. Liparidae | 0.800 | 11.92 | ||
| Сем. Cottidae | 1.727±0.235 | 30.49±4.15 | ||
| Подотряд Scorpaenoidei | 1.898±0.345 | 34.79±6.32 | ||
| Сем. Scorpaenidae | 1.267 | 25.11 | ||
| Сем. Triglidae | 2.739 | 47.70 | ||
| Отряд Pleuronectiformes | 1.762±0.182 | 29.49±3.05 | ||
| Подотряд Pleuronectoidei | 1.777±0.199 | 29.11±3.26 | ||
| Подотряд Soleoidei | 1.691 | 31.27 | ||
| Сем. Cynoglossidae | 0.648 | 16.04 | ||
| Сем. Soleidae | 2.212 | 38.89 | ||
| Отряд Mugiliformes | 1.411 | 29.95 | ||
| Отряд Gasterosteiformes | 1.739±0.233 | 31.60±4.23 | ||
| Подотряд Syngnathoidei | 1.480±0.224 | 28.87±4.37 | ||
| Сем. Syngnathidae | 1.437±0.220 | 27.55±4.22 | ||
| Сем. Carangidae | 1.671 | 34.79 | ||
| Подотряд Gasterosteoidei | 2.311 | 37.62 | ||
| Отряд Perciformes | 1.884±0.106 | 33.09±1.86 | ||
| Подотряд Zoarcoidei | 1.115±0.243 | 16.61±3.62 | ||
| Подотряд Anabantoidei | 0.862 | 21.31 | ||
| Сем. Anabanyidae | 0.831 | 20.55 | ||
| Сем. Chanidae | 0.892 | 22.07 |
| Подотряд Trachinoidei | 1.733 | 25.82 | ||
| Подотряд Gobioidei | 1.305±0.194 | 27.30±4.06 | ||
| Сем. Gobiidae | 1.148±0.167 | 25.00±3.34 | ||
| Сем. Periophthalidae | 2.324 | 42.26 | ||
| Подотряд Callionymoidei | 1.914 | 28.51 | ||
| Подотряд Percoidei | 1.742±0.210 | 31.73±3.83 | ||
| Сем. Cepolidae | 0.326 | 6.33 | ||
| Сем. Grammistidae | 0.496 | 12.27 | ||
| Сем. Kuhliidae | 0.586 | 14.49 | ||
| Сем. Centrarchidae | 1.055±0.155 | 17.95±2.64 | ||
| Сем. Cichlidae | 0.934±0.091 | 23.11 | ||
| Сем. Serranidae | 1.302 | 32.19 | ||
| Сем. Percidae | 2.232±0.176 | 33.47±2.06 | ||
| Сем. Centracanthidae | 1.705±0.195 | 33.78±3.86 | ||
| Сем. Embiotocidae | 2.289 | 34.11 | ||
| Сем. Mullidae | 1.643 | 40.63 | ||
| Сем. Girellidae | 1.742 | 43.08 | ||
| Сем. Percichthydae | 3.424±0.375 | 51.02±5.59 | ||
| Сем. Pomadagyidae | 2.087 | 51.62 | ||
| Сем. Sparidae | 2.891±0.490 | 58.31±9.88 | ||
| Сем. Sciaenidae | 3.033 | 75.02 | ||
| Подотряд Labroidei | 1.423 | 31.94 | ||
| Подотряд Blennioidei | 1.705 | 35.89 | ||
| Подотряд Notothenioidei | 3.443±0.372 | 51.31±5.54 | ||
| Сем. Bathydraconidae | 2.490 | 37.10 | ||
| Сем. Chaenichthyidae | 2.748±0.477 | 40.94±7.11 | ||
| Сем. Nototheniidae | 3.846±0.504 | 57.31±7.51 | ||
| Подотряд Echeneoidei | 2.885 | 71.35 | ||
| Надотряд Parapercomorpha | 2.114±0.336 | 33.00±5.25 | ||
| Отряд Gadiformes | 2.114±0.336 | 33.00±5.25 | ||
| Подотряд Ophidoidei | 0.302 | 7.46 | ||
| Подотряд Gadoidei | 2.316±0.300 | 35.83±4.64 | ||
| Сем. Macrouridae | 0.359 | 8.87 | ||
| Сем. Gadidae | 2.560±0.198 | 39.20±5.48 | ||
| Надотряд Osteoglossomorpha | 1.416 | 35.03 | ||
| Надотряд Clupeomorpha | 2.378±0.090 | 35.79±1.35 | ||
| Отряд Salmoniformes | 2.331±0.082 | 34.72±1.22 | ||
| Подотряд Osmeroidei | 1.813 | 27.01 | ||
| Подотряд Esocoidei | 2.011±0.194 | 29.97±2.89 | ||
| Подотряд Salmonoidei | 2.390±0.089 | 35.27±1.31 | ||
| Сем. Salmonidae | 2.313±0.089 | 34.46±1.33 | ||
| Сем. Coregonidae | 2.803±0.324 | 41.77±4.83 | ||
| Сем. Thymallidae | 3.424 | 51.01 | ||
| Отряд Gonorhynchiformes | 2.131 | 52.70 | ||
| Отряд Clupeiformes | 3.779 | 56.31 |
Рассчитанные по собранным нами литературным данным (Зотин, Зотин, 1999) коэффициенты а и Сr (при k=0.75) представлены в табл. 34(3). Классификация костных рыб в этой таблице дана по Рассу (1971) и Решетникову и др. (1989), но расположение отдельных таксонов приведено в соответствии с величиной критерия упорядоченности. Расположение отрядов в табл. 34(3) отличается от приводимого Озернюком и Булгаковой (1997), хотя принципы заложенные в основу этой и нашей работы почти одинаковые. Это показывает насколько еще ненадежно, при имеющихся в настоящее время в литературе данных о стандартном обмене рыб, использование коэффициента а и критерия упорядоченности для построения макросистематики рыб. И все же расположение подклассов в классе Osteichthyes можно представить согласно данным, приводимым в табл.34 (3) в следующем виде:
Sarcopterygii -> Actinopterygii -> Teleostei,
а предварительное расположение надотрядов в подклассе Teleostei в виде:
Дата публикования: 2014-11-04; Прочитано: 418 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
