 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Разложение лигнина
|
|
Лигноцеллюлоза составляет почти половину годичной продукции наземной растительности. Лигноцеллюлоза составляет 90-98% древесины. По обилию лигнин стоит на втором месте после целлюлозы. Лигнин, составляя углеродный скелет растений, трудно разлагается и поэтому представляет один из важных компонентов резервуаров углерода с длительным временем пребывания, и его разложение представляет узкое место в круговороте углерода в наземных системах. В лигноцеллюлозе лигнин связан с гемицеллюлозой, в то время как его связь с целлюлозными фибриллами скорее физическая, чем химическая. Поэтому пути деградации лигноцеллюлозы могут идти по двум независимым маршрутам: разложение целлюлозы группировкой “бурой гнили” или же разложение гемицеллюлозно-лигнинового комплекса организмами “белой гнили”, наиболее известным примером которых может служить опенок Armillariella, а наиболее частым лабораторным объектом - Phanerochaete. Грибы, растущие на лигноцеллюлозе представляют наиболее разнообразную и многочисленную группировку сапрофитных грибов, макро- и микромицетов. Позднее появление лигноцеллюлозы в истории биоты в фанерозое, связанное с появлением наземного растительного покрова, предполагает существование в условиях ранее созданной аэробной атмосферы и развитие грибов как наземной группировки органотрофных ксерофильных аэробных организмов, развивающихся в виде мицелия на твердом субстрате. Разложение лигнина происходит за счет аэробных процессов с участием активного кислорода, необходимого для разрыва связей между фенилпропановыми частями молекулы лигнина. Предполагается, что ароматические соединения, образующиеся при разложении лигнина, лишь замедленно разлагаются в аэробных условиях и являются исходным материалом для конденсации в молекулы гуминовых соединений, возможно, с участием микробных внеклеточных полимераз в биологически опосредованных реакциях.
Обычно считается, что в анаэробных условиях лигнин инертен, но это не относится к олигомерным продуктам его разложения, которые могут метаболизироватъся бактериями, атакующими в анаэробных условиях фенолы (Schink et al., 2000).
Анаэробное разложение полученных из лигнина соединений может осуществляться в условиях денитрификации, сульфидогенеза, метаногенеза и подробно исследовано в последние десятилетия Г. Фуксом, Б.Шинком после пионерских работ Г. Зейкуса по разложению лигнина бактериями. Эти исследования особенно усилились в связи с изучением деградации галогенизированной ароматики, поступающей от ксенобиотиков в виде загрязнения.
Общая схема разложения древесины включает разделение ее на компоненты, которые разлагаются разными группировками микроорганизмов.
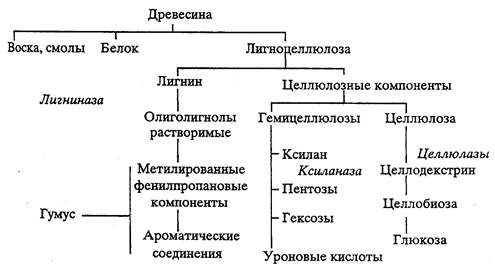
Дезинтеграция лигнина включает несколько этапов. На первом из них происходит образование лигнолов, растворимых остатков лигнина, представляющих случайное собрание гидрофильных молекул умеренного молекулярного веса. Образуются эти молекулы в результате действия “лигниназ” - ферментов, продуцирующих при соокислении органического вещества продукты неполного восстановления О2 - перекиси и даже радикалы. Перекисный механизм разложения лигнина приводит к хаотическому разрыву связей, но образующиеся осколки оказываются, более доступны для микроорганизмов, и поэтому их концентрация в системе определяется динамическим равновесием между продукцией и разложением. Следующий этап разложения приводит к образованию крупных молекул ароматических кислот: феруловой, сирингиловой, циннамовой, ванилиновой. В дополнение к ним имеется множество осколков структуры в виде циклических соединений, особенно фенолов, катехинов. В результате деметилирования поступают и С-1 соединения. Размеры молекул этих субстратов позволяют осуществлять их осмотрофное использование с поступлением в клетку. Широкий набор субстратов допускает две возможности: действия микроорганизмов- универсалистов, использующих все эти соединения внутри клетки, или же действия микробов-специалистов в составе сообщества. Обе эти возможности осуществляются. Многие грибы, растущие на древесине, оказываются в достаточной степени универсалистами.
Разложение циклических соединений аэробными микроорганизмами внутри физической структуры разлагаемых растительных остатков неизбежно приводит к дефициту кислорода, которому способствует влажность. Аноксические условия способствуют развитию бактериальной микрофлоры, способной к разложению циклических соединений без участия кислорода и без доступа внешних окислителей, как нитрат или сульфат. В этих условиях, помимо способности к брожению, оказывается важным использование органических веществ как акцепторов электрона. В качестве примера таких процессов можно назвать фумаровое дыхание и, что особенно интересно, открытое недавно хиноновое дыхание. В какой мере хиноны могут служить диффундирующим переносчиком между восстановленной частью структуры и поверхностью, где они могут окислиться и вернуться обратно, а не быть съеденными аэробами, неясно.
Разложение лигнина, как было упомянуто выше, ведет к образованию веществ разной молекулярной массы. Среди них доминируют циклические соединения. Предполагается, что эти соединения, обладающие реакционно-способными функциональными группами, могут реагировать между собой, объединяясь в крупные макромолекулярные соединения путем химической конденсации, возможно, под действием неспецифически активирующих процесс ферментов. Происходит так называемая поликонденсация с синтезом гумусовых; веществ. В нее вовлекается ряд веществ микробного происхождения. Эта схема описывает представления о происхождении гуминовых соединений из продуктов разложения лигнина. При этом исходные химические структуры лигнина в гумусе уже перестают быть опознаваемыми вследствие перестройки на уровне относительно мелких молекул, вплоть до фенолов.
Описываемая последовательность событий в главном для наземных биоценозов маршруте превращений органического вещества вряд ли может быть прослежена по судьбе химических структур ввиду их разнообразия. В общих чертах предполагается первоначальная деполимеризация лигнина от молекулярных масс порядка мегадальтон к их осколкам размером в тысячи-сотни дальтон с последующим укрупнением в макромолекулы порядка десятков-сотен тысяч дальтон. Поскольку сам лигнин подвергается при своем биосинтезе процессу ферментативной конденсации из фенольных предшественников, то представляется вероятной иммобилизация аналогичным образом органического углерода путем образования гумусовых соединений, но из гораздо большего разнообразия предшественников и не под таким строгим контролем, как в клетке растения. При этом прямое участие биоты уже ограничено, и это лишает процесс специфичности.
В масштабах России процесс вероятного синтеза гумуса из лигнина как основного предшественника количественно согласуется с поступлением лигноцеллюлозы, которое с избытком перекрывает оценку синтеза гумуса порядка 200 Мт С/год, если считать время пребывания углерода в гумусе около тысячи лет. Дополнительным источником вещества в гумусе, и особенно азотистых компонентов, следует считать деградацию мицелия грибов как вторичных продуцентов мортмассы в трофической системе. Отсюда следует, что в образовании гуминовых соединений лигнин в первую очередь играет роль как поставщик циклических соединений, но он является не единственным предшественником, а лишь важным составным компонентом.
Лигноцеллюлоза представляет важную составную часть торфа - не полностью разложенных гумифицированных растительных остатков. Микробные группировки, участвующие в процессе деградации лигноцеллюлозы, следующие:
1 Лигнолитический маршрут:
Дата публикования: 2015-10-09; Прочитано: 1701 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
