 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Запасы органического углерода в слое 0-100 см почв и торфов России
|
|
[по: Орлов и др., 1996]
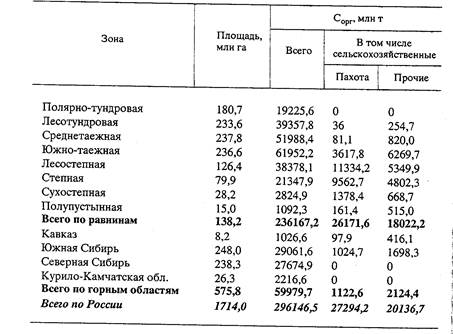
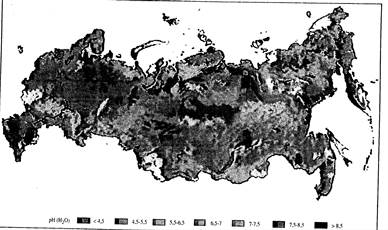
Рис. 7.2. Кислотность верхнего горизонта почвы [по: Stolbovoi, McCallum, 2002]
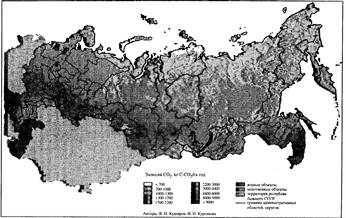
Рис- 7.3. Годовая эмиссия углекислого газа из почв России [по: Кудеяров, 1999]
Рис. 7.4. Картограмма запасов гумуса в почвах России, т/га [по: Орлов, 1999]
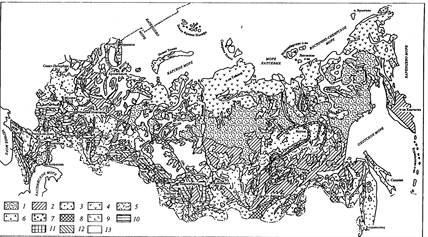
Рис. 7.5. Гидролиз целлюлозы [по: Tomme et al., 1995]

I - ТАК; 2 - ксилогликан; 3 - поперечные сшивки; 4 - фенольные сшивки
Модель фрагмента первичной клеточной стенки травянистых растений иллюстрирует трудности, с которыми сталкиваются микроорганизмы при гидролизе растительных остатков. Модель основания на анализе колеоптиля кукурузы и др. однодольных. Показан один слой микрофибрилл целлюлозы непосредственно после деления клеток; множество таких слоев срастаются, образуя клеточную стенку. Микрофибриллы связываются глюкуроноарабиноксиланами (ГАК), скрепляющимися с микрофибриллами фенольными сшивками. В стенках многих других высших растений во взаимосвязях участвуют молекулы ксилогликана, присоединяющиеся водородными мостиками к поверхности микрофибрилл
Скорость процесса разложения приводит к тому, что развитие этой группировки зимогенных бактерий носит эфемерный характер. Из легкоразлагаемых углеводов первое место занимает крахмал как запасное вещество с группировкой аэробных и анаэробных амилолитических бактерий. Более устойчивы к разложению структурные компоненты растительной ткани, как пектин. Имеется специализированная группа бактерий, осуществляющая мацерацию растительной ткани и представленная бактериями мочки льна. Защитные слизи и камеди разлагаются медленнее, но и здесь действует специфическая группировка пуллуланолитических бактерий с гидролитическим ферментом пуллуланазой. В слизетечениях развивается Leuconostoc, в свою очередь образующий слизь.
Растительные оболочки (рис. 7.5) содержат структуры, составленные из: а) микрофибрилл целлюлозы с кристаллическими и аморфными участками, последние легче гидролизуются; б) гемицеллюлозы с ксиланами и глюкоманнанами, разделяющими целлюлозные фибриллы; в) ксилана из ксилозной цепи с глюкуроновой кислотой и арабинозой, этерифицированной феноловыми кислотами, в свою очередь связанной с лигнином ковалентными связями; г) лигнина, представляющего нерегулярную сеть из ароматических компонентов, как фенилпропан, и устойчивого к микробному разложению; д) пектина, представляющего слизи срединной пластинки, из полиуроновых кислот, например галактуроновой кислоты, этери- фицированных метанолом. Метанол отщепляется метилэстеразой. Метальные группы пектина и лигнина служат источником С-1 соединений для метилотрофов. В водорослях аналогом пектина является альгиновая кислота. Водоросли не содержат лигнина, являющегося характерным веществом для устойчивого углеродного скелета наземной растительности.
В море целлюлоза не является доминирующим структурным полисахаридом. Здесь ее заменяет хитин скелета морских беспозвоночных, по обилию на Земле представляющий второй или третий полисахарид. На суше помимо беспозвоночных источником хитина служит клеточная стенка грибов. Гидролизуется хитин экзоферментом хитиназой. Хитин гидролизуют микофильные грибы, а также широкий набор бактерий как аэробных, в том числе актиномицеты, так и анаэробных. Среди бактерий следует отметить помимо стреп- томицетов аэробные псевдомонады, цитофаги, флавобактерии, артробактеры, бациллы, Lysobacter (лизирующий структурно сходный с хитином муреин бактерий) и анаэробные клостридии, Halanaerobacter. Характерными потребителями ацетилглюкозамина служат планктомицеты.
Как уже отмечалось, полиглюкоза представлена растворимыми легкодоступными соединениями, например крахмалом, и труднодоступными нерастворимыми компонентами оболочек, прежде всего целлюлозой. Крахмал содержит неразветвленную амилозу с а-1,4- и разветвленный амилопектин с а-1,6-связями. Это легкодоступное соединение, гидролизуемое многими микроорганизмами, образующими амилазу. Довольно легко используется и пектин, для которого характерными являются Erwinia, Pectinatus и организмы мочки льна “Bacillus felsineus". Осуществляют гидролиз пектина пектиназы с характерным побочным продуктом метанолом.
Целлюлоза имеет молекулярный вес около миллиона, Р-1,4- связь в цепочке и водородные связи между параллельными цепочками. Гидролизуется целлюлоза целлюлазным комплексом, который осуществляет сначала разрушение кристаллической структуры, затем деполимеризацию цепочки на дисахарид целлобиозу экзоглюканазой или целлодекстрины эндоглюканазой. Целлобиоза гидролизуется целлобиазой с образованием глюкозы, которая может транспортароваться в клетку. Целлюлозолитические организмы представляют важнейшую группу, открывающую основной маршрут деструкции; органического углерода на Земле. Разложение целлюлозы оказывается лимитирующим звеном в круговороте углерода в наземных экосистемах и подробнее рассмотрено ниже.
Гемицеллюлозы представляют сборную группу полимеров гексоз, пентоз, уроновых кислот. Обычно доминируют ксилоза и малноза. Гемицеллюлозы сами по себе легко гидролизуются, но в природе они обычно связаны с другими полимерами, и их разложение затруднено.
Ксилан представляет гетерополимер из цепи ксилозы с арабинозой, глюкозой, галактозой, и другими сахарами как боковыми цепями. Типичной группировкой бактерий, деградирующих ксилан, являются клостридии с низким Г + Ц, как Clostridium polysaccharolyticum, С. cellulovorans, С. xylanolyticum, С. xylanovorans, а также представители родов Eubacterium, Fibrobacter, Butyrivibrio, Selenomonas, Thermotoga. Он относительно легко сбраживается анаэробными opганизмами, которые используют широкий набор олигосахаридов, образующихся после гидролиза ксиланазами.
Гидролиз полиглюкозы и вообще полисахаридов микроорганизмами обычно подавляется при высоком содержании продуктов гидролиза в среде. Отсюда этот маршрут специально обусловлен взаимодействием гидролитиков с диссипотрофами. Хитин состоит из полимера М-ацетил-Б-глюкозамина. Как уже отмечалось, это основной компонент клеточной стенки грибов. Хитин ассоциируется с другими полимерами. Его разложение в большой степени зависит от физического состояния и доступности; поверхности для действия хитиназы и других гидролаз. Темноокрашенные пигменты меланины обусловлены связью сахаров с фенольньши соединениями. Аминосахара, включая ацетилглюкозамин и ацетилмурамовую кислоту, присутствуют и в стенке бактерий. Разложение стенок грибов идет относительно быстро и легко под действием, как бактерий, так и микофильных грибов, которые доминируют в более кислых условиях. Однако в целом стенки и мембраны представляют относительно устойчивый компонент микробной мортмассы. Считается, что аминосоединения микробных стенок служат предшественниками азота в гумусе.
Лигнин представляет условие появления наземной растительности. Это разветвленный полимер фенил-пропаноида. Лигнин образуется химической поликонденсацией фенола под действием свободного радикала и не имеет закономерного строения. Он связывается с целлюлозой и гемицеллюлозами, инкрустируя их. В результате получается устойчивое, вследствие своей гетерогенности и неупорядоченности, соединение, которое не поддается прямому энзиматическому гидролизу. Деградация лигнина осуществляется почти исключительно аэробными микроорганизмами. Исключение составляет микробное сообщество пищеварительного тракта термитов. Деградация лигнина почти недоступна прокариотам, и о возможности их участия в деградации меченого по углероду лигнина идут споры. Во всяком случае, процесс если и есть, то очень слабый.
Деградацию лигнина осуществляют грибы “белой гнили” Polyporus, Pleurotus, Phanerochaete. Обычно для начала деградации необходимо наличие легкодоступного субстрата дыхания. Напротив, грибы “бурой гнили” разлагают углеводные компоненты и оставляют фенольную часть, которая окисляется и темнеет. Продукты деградации лигнина, в том числе ароматические соединения, доступны как аэробным, так и анаэробным бактериям. Проведено очень большое число работ по разложению модельных компонентов лигнина, таких, как кофеиновая кислота. Особенно быстро разложение идет в аэробно/анаэробном сообществе.
Частицы органического вещества представлены в основном оболочками отмерших организмов и состоят из нерастворимых полимеров. Они входят в относительно медленно метаболизируемую органику, которая становится доступной для использования осмотрофными организмами только после гидролиза до олигомеров. Гидролиз осуществляется вне клетки, и поэтому гидролитические экзоферменты должны быть устойчивы и активны в условиях среды. Для экстремофильных гидролитических организмов эта необходимость накладывает дополнительное ограничение: кроме поддержания внутриклеточного гомеостаза клетки должны вырабатывать устойчивые и активные белки-экзоферменты. Гидролитические экзоферменты необходимы группам гидролитических организмов, использующим целлюлозу и другие полисахариды, хитин, нерастворимые белки, липиды.
Гидролитические экзоферменты бактерий могут либо располагаться на внешней поверхности клетки, либо секретироваться в среду. Во втором случае продукты гидролиза могут стать доступными не только продуценту, но и случайным организмам в окружении. Типичную группировку такого типа и составляют диссипотрофы. Расположение гидролитических ферментов на клеточной поверхности предполагает их заякоривание в липидной части поверхностных структур, что, в свою очередь, предполагает необходимость тесного физического контакта клетки с гидролизуемым материалом. Отсюда физическая организация взаимодействия клетки с гидролизуемым материалом приобретает первостепенное значение. С одной стороны, организм должен выработать механизмы адгезии к гидролизуемому субстрату, а с другой - сохранить способность перемещаться с уже выработанных участков на свежие. Это требование может быть выполнено для организмов, образующих подвижные клетки для расселения, или же, что еще выгоднее, для организмов со скользящим типом движения, расползающихся по поверхности субстрата. Образование слизистой капсулы на поверхности гидролизуемого сложного субстрата создает возможности для кооперативного действия сообщества разнородных организмов, воздействующих на разные компоненты сложного нерастворимого субстрата. Регуляция синтеза экзоферментов подчиняется общим правилам катаболитной репрессии и индукции. Считается, что обычно организмы не способны синтезировать экзоферменты в присутствии растворимых легкоусваиваемых веществ, в том числе продуктов гидролиза.
Альтернативой гидролитическому пути разложения полимеров служит расщепление взвешенного органического вещества с нерегулярным строением под действием активного кислорода. Примером такого способа деградации служит процесс разложения лигнина экзоферментами, использующими как источник активного кислорода Н202, образующуюся при дыхании клетки, он носит общее название соокисления труднодоступных субстратов. Очевидно, что этот способ деградации доступен только аэробам. Он распространен преимущественно у грибов. Разложение негидролизуемого полимерного вещества анаэробами происходит медленно и в водных экосистемах часто требует участия сообщества микроорганизмов с сульфатредукторами как терминальной группировкой. В наземных условиях сульфидогенез ограничен, и основным путем разложения в аноксической обстановке служит метаногенез.
Суммарный процесс разложения целлюлозы в почве оценивается аппликационным методом по И. Вострову: в почвенный профиль закладывается ткань и после инкубации выявляется область разрушения, связанная с определенным слоем почвы.
Дата публикования: 2015-10-09; Прочитано: 714 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
