 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Энергетический метаболизм
|
|
Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий реакции, связанные с получением энергии в форме АТФ, - это реакции окисления- восстановления, сопряженные с реакциями фосфорилирования. Окисленный в этих реакциях углерод выделяется клеткой в виде СО2. Для удаления отщепившегося в этих реакциях водорода, который находится в форме восстановленного НАД, разные бактерии используют различные возможности в зависимости от конечного акцептора водорода (или электронов, что является эквивалентным понятием). В зависимости от способа получения энергии у бактерий имеется несколько типов метаболизма: окислительный, или дыхание; бродильный, или ферментативный; смешанный. Тип метаболизма определяет не только реакции, в результате которых образуется АТФ, но и конечные продукты этих реакций, которые используются при идентификации бактерий, а также условия культивирования бактерий.
При использовании в качестве источника углерода и энергии глюкозы или других гексоз начальные этапы окисления глюкозы являются общими, как при оксидативном, так и при бродильном метаболизме. К ним относятся пути превращения глюкозы в пируват (при использовании в качестве источника энергии отличных от глюкозы гексоз, или дисахаридов, они в результате химических превращений вступают в цепь реакций, превращающих глюкозу в пируват). Пируват, образовавшийся при расщеплении глюкозы, превращается при участии кофакторов в активированную уксусную кислоту или ацетилкоэнзим А. Последний окисляется в СО2 с отщеплением водорода в цикле трикарбоновых кислот.
Цикл трикарбоновых кислот не только выполняет функцию конечного окисления питательных веществ, но и обеспечивает процессы биосинтеза многочисленными предшественниками: пируват α-кетоглутаровая, щавелевая и янтарные кислоты - для синтеза аминокислот, щавелевоуксусная - для синтеза пиримидиновых нуклеотидов, малонат - для синтеза аминокислот, пиримидиновых нуклеотидов и жиров (рис. 4.2).
Окислительный метаболизм. Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания. Дыхание - процесс получения энергии в реакциях окисления-восстановления, сопряженных с реакциями окислительного фосфорилирования, при котором донорами электронов могут быть органические (у органотрофов) и неорганические (у литотрофов) соединения, а акцептором - только неорганические соединения.
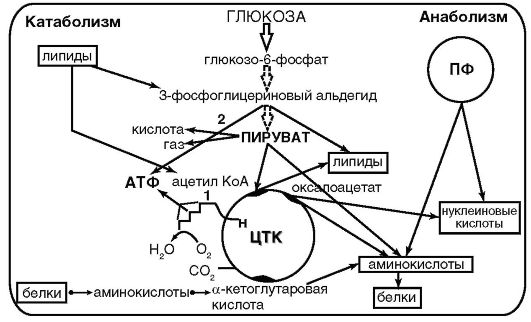
Рис. 4.2. Схема обмена веществ у бактерий
У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода [Н+]) является молекулярный кислород. В этом случае пируват полностью окисляется в цикле трикарбоновых кислот до СО2. Цикл трикарбоновых кислот выполняет функции поставщика как предшественников для биосинтетических процессов, так и атомов водорода, который в форме восстановленного НАД переносится на молекулярный кислород через серию переносчиков, обладающих сложной структурно оформленной мультиферментной системой - дыхательной цепью. Дыхательная цепь у бактерий локализована в ЦПМ и во внутриклеточных мембранных структурах.
Электрохимическую энергию бактерии получают в процессе переноса электронов по окислительно-восстановительным цепям в мембране, в результате чего происходит неравномерное распределение Н+ по обеим ее сторонам. Переносчики электронов располагаются в мембране таким образом, что во внешней среде происходит накопление ионов водорода (при этом возникает подкисление среды), а в цитоплазме их число уменьшается, что сопровождается подщелачиванием среды. Неравномерное распределение положительно заряженных протонов (большее число на наружной и меньшее на внутренней поверхности плазматической мембраны) приводит к формированию расположенного поперек мембраны электрического поля, мембранного потенциала. В результате при переносе электронов возникает трансмембранный электрохимический градиент ионов водорода, обозначаемый символом ΔμΗ+ и измеряемый в вольтах. Энергия мембранного потенциала используется для синтеза локализованной в мембране АТФазой АТФ.
Энергия в форме ΔμΗ+ не теряется при ее запасании и может образовываться и потребляться клеткой в условиях, когда невозможен синтез АТФ. В последние годы показано, что аналогичным образом перераспределяются и атомы Na+ с образованием энергии, обозначаемой как ΔμNa+. Данные формы энергии тратятся преимущественно на движение бактерий (у подвижных форм) и транспорт веществ в клетку и из нее.
Типичная цепь выглядит следующим образом: ЦТК → НАД(Н2) → флавопротеид → хинон → цитохромы: b c a → О2.
Конечным этапом переноса электронов (протонов) по дыхательной цепи является восстановление цитохромов а + а3 (цитохромоксидазы). Цитохромоксидаза является конечной оксидазой, передающей электроны на кислород. Образующиеся при окислении ФАД или хинонов протоны связываются ионами О2- с образованием воды.
В то время как у эукариотов ферменты дыхательной цепи имеют относительно постоянный состав, у бактерий встречаются вариации в составе дыхательной цепи. У некоторых бактерий цитохромы отсутствуют и при контакте с кислородом происходит непосредственный перенос водорода на кислород с помощью флавопротеидов, конечным продуктом при этом оказывается перекись водорода (Н2О2).
Помимо углеводов, прокариоты способны использовать другие органические соединения, в частности белки, в качестве источника энергии, окисляя их полностью до СО2 и Н2О.
Аминокислоты и белки также могут выступать в качестве энергетических ресурсов. Их использование связано в первую очередь с определенными ферментативными преобразованиями подготовительного характера. Белки вначале вне клетки расщепляются протеолитическими ферментами на пептиды, которые поглощаются клеткой и расщепляются внутриклеточными пептидазами до аминокислот. Аминокислоты могут использоваться в конструктивном метаболизме, а могут у аммонифицирующих бактерий служить основным материалом в энергетических процессах при окислительном дезаминировании, в результате которого происходят выделение аммиака и превращение аминокислоты в кетокислоту, которая через цикл трикарбоновых кислот вступает в конструктивный метаболизм.
Процесс аммонификации известен как гниение, при этом происходит накопление продуктов, обладающих неприятным специфическим запахом образующихся при этом первичных аминов. Гнилостные бактерии осуществляют минерализацию белка, разлагая его до СО2, NH3, H2S. К гнилостным бактериям относятся Proteus, Pseudomonas, Bacillus cereus.
Анаэробное дыхание. Некоторые бактерии обладают способностью использовать в анаэробных условиях нитрат как конечный акцептор водорода. Восстановление нитрата может происходить двумя путями: аммонификацией, при которой нитрат превращается в аммиак, и денитрофикацией, при которой происходит восстановление нитрата до молекулярного азота или закиси азота. Этот процесс связан с деятельностью фермента нитратредуктазы.
Сульфатное дыхание. Использовать сульфат как конечный акцептор водорода при анаэробном дыхании способна лишь небольшая группа бактерий, включающая только два рода: Desulfovibrio, Desulfotomaculum. Эти бактерии являются строгими анаэробами, они обитают в сероводородном иле и не имеют значения в медицинской микробиологии. Они способны использовать в качестве донора электронов молекулярный водород, поэтому их относят к хемолитотрофам. Этим бактериям принадлежит ведущая роль в образовании сероводорода в природе.
Дата публикования: 2015-10-09; Прочитано: 2155 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
