 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Химический состав бактериальной клетки
|
|
Бактериальная клетка на 80-90% состоит из воды и только 10% приходится на долю сухого вещества. Вода в клетке находится в свободном или связанном состоянии. Она выполняет механическую роль в обеспечении тургора, участвует в гидролитических реакциях. Удаление воды из клетки путем высушивания приводит к приостановке процессов метаболизма, прекращению размножения, а для многих микроорганизмов губительно. В то же время особый способ высушивания микроорганизмов в вакууме из замороженного состояния (лиофилизация) обеспечивает сохранение жизнеспособности большинства микроорганизмов. Лиофилизация используется для приготовления проб, пригодных для длительного хранения.
В сухом веществе бактерий 52% составляют белки, 17% - углеводы, 9% - липиды, 16% - РНК, 3% - ДНК и 3% - минеральные вещества.
Белки являются ферментами, а также составной частью клетки, входят в состав цитоплазматической мембраны (ЦПМ) и ее производных, клеточной стенки, жгутиков, спор и некоторых капсул. Некоторые бактериальные белки являются антигенами и токсинами бактерий. В состав белков бактерий входят отсутствующие у человека D-аминокислоты, а также диаминопимелиновая кислота.
Углеводы представлены в бактериальной клетке в виде моно-, ди-, олигосахаров и полисахаридов, а также входят в состав комплексных соединений с белками, липидами и другими соединениями. Полисахариды входят в состав некоторых капсул, клеточной стенки; крахмал и гликоген являются запасными питательными веществами. Некоторые полисахариды принимают участие в формировании антигенов.
Липиды или жиры входят в состав ЦПМ и ее производных, клеточной стенки грамотрицательных бактерий, а также служат запасными веществами, входят в состав эндотоксина грамотрицательных бактерий, в составе ЛПС формируют антигены. В бактериальных жирах преобладают длинноцепочечные (С14-С18) насыщенные жирные кислоты и ненасыщенные жирные кислоты, содержащие одну двойную связь. Сложные липиды представлены фосфатидилинозитом, фосфатидилглицерином и фосфатидилэтаноламином. У некоторых бактерий в клетке находятся воски, эфиры миколовой кислоты. Микоплазмы - единственные представители царства Procaryotae, имеющие в составе ЦПМ стеролы. Остальные бактерии в составе ЦПМ и ее производных не имеют стеролов.
В бактериальной клетке присутствуют все типы РНК: иРНК, транспортная РНК (тРНК), рРНК, менее известная антисенс РНК (асРНК). Молекулы асРНК пока не обнаружены в клетках эукариот. Информация об асРНК записана в хромосоме, в так называемых антисенс-генах. АсРНК принимает активное участие в регуляции различных клеточных процессов, в том числе репликации ДНК бактерий, вирусов, плазмид и танспозонов. асРНК представляет собой короткую молекулу, комплементарную определенному участку иРНК, и, соединяясь с ней, блокирует процесс синтеза белка. При этом в клетке подобные комплексы могут накапливаться, и при диссоциации асРНК и иРНК одновременно начинается синтез белка на большом числе однотипньгх матриц. Искусственные молекулы асРНК пытаются использовать для борьбы с бактериями за счет угнетения ими синтеза в клетке определенных жизненно важных белков.
Пуриновые и пиримидиновые нуклеотиды - это те строительные блоки, из которых синтезируются нуклеиновые кислоты. Кроме того, пуриновые и пиримидиновые нуклеотиды входят в состав многих коферментов и служат для активации и переноса аминокислот, моносахаров, органических кислот.
ДНК выполняет в бактериальной клетке наследственную функцию. Молекула ДНК построена из двух полинуклеотидных цепочек. Каждый нуклеотид состоит из азотистого основания, сахара дезоксирибозы и фосфатной группы (рис. 4.1, б). Азотистые основания представлены пуринами (аденин, гуанин) и пиримидинами (тимин, цитозин). Каждый нуклеотид обладает полярностью. У него имеется дезоксирибозный З'-конец и фосфатный 5'-конец. Нуклеотиды соединяются в полинуклеотидную цепочку посредством фосфодиэфирных связей между 5'-концом одного нуклеотида и З'-концом другого (рис. 4.1, а). Соединение цепей обеспечивается водородными связями между комплементарными азотистыми основаниями: аденина с тимином, гуанина с цитозином.
Нуклеотидные цепи антипараллельны: на каждом из концов линейной молекулы ДНК расположены 5'-конец одной цепи и З'-конец другой цепи. Процентное содержание ГЦ-пар в ДНК определяет степень родства между бактериями и используется при определении таксономического положения бактерий.
Минеральные вещества обнаруживаются в золе, полученной после сжигания клеток. В большом количестве представлены N, S, Р, Са, К, Mg, Fe, Mn, а также микроэлементы Zn, Cu, Co, Ва.
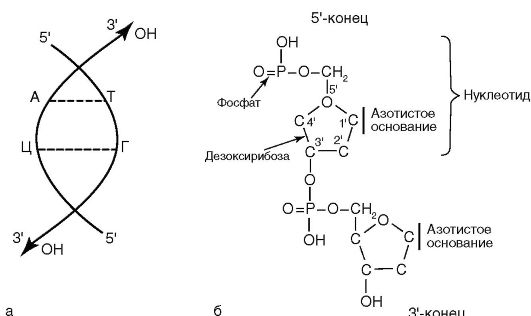
Рис. 4.1. Строение ДНК и ее элементов (объяснение в тексте)
Азот входит в состав белков, нуклеотидов, коферментов. Сера входит в виде сульфгидрильных групп в структуру белков. Фосфор в виде фосфатов представлен в нуклеиновых кислотах, АТФ, коферментах. В качестве активаторов ферментов используются ионы Mg, Fe, Mn. Ионы К и Mg необходимы для активации рибосом. Са является составной частью клеточной стенки грамположительных бактерий. У многих бактерий имеются сидерохромы, которые обеспечивают транспортировку ионов Fe внутрь клетки в виде растворимых комплексных соединений.
Конструктивный метаболизм. Основные органические компоненты бактериальной клетки, как уже было отмечено, синтезируются в реакциях полимеризации из строительных блоков: аминокислот, фосфатов, сахаров, пуриновых и пиримидиновых оснований, органических кислот. Поставщиками этих строительных блоков являются промежуточные продукты основных путей энергетического метаболизма (см. рис. 3.2). Среди бактерий выделяется группа, называющаяся прототрофами, которые способны синтезировать все компоненты клетки из одного источника углерода и энергии. Если бактерии теряют способность образовывать какое-либо жизненно важное вещество (аминокислоту, витамин, фактор роста и др.), участвующее в биосинтетических процессах, то для их роста и размножения требуется его поступление в готовом виде. Такие вещества называют фактором роста, а бактерии, возникшие, как правило, в результате мутаций, - ауксотрофами.
Факторами роста являются аминокислоты, пуриновые и пиримидиновые основания, витамины, которые входят в состав простетических групп коферментов.
Биосинтез аминокислот и синтез белка. Большинство бактерий обладают способностью синтезировать все 20 аминокислот, из которых состоят белки. Белки в бактериальной клетке выполняют ферментативную функцию, а также являются составной частью структурных образований клетки: ЦПМ и ее производных, клеточной стенки, жгутиков, капсулы и спор у некоторых бактерий.
Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные продукты фруктозодифосфатного (ФДФ) и пентозофосфатного (ПФ) путей, цикл трикарбоновых кислот: пируват, кетоглутаровая кислота, оксалоацетат, фумарат, эритрозо-4-фосфат, рибозо-4-фосфат. Аминогруппы вводятся в результате непосредственного аминирования или переаминирования. Перевод неорганического азота в органические соединения происходит всегда через аммиак. Нитраты и нитриты и молекулярный азот предварительно восстанавливаются в аммиак и только лишь после этого включаются в состав органических соединений. В результате прямого аминирования образуются лишь L-аланин, L-аспартат, L-глутамат и L-глугамин. Все остальные аминокислоты получают свою аминогруппу в результате переаминирования с одной из первичных аминокислот. Синтез белка осуществляется у бактерий так же, как в клетках эукариот.
Синтез белка происходит на рибосомах и обычно подразделяется на три процесса: инициацию, элонгацию и терминацию. Инициация синтеза белка заключается в связывании мет-тРНК с малой субъединицей рибосомы с последующим встраиванием инициирующего кодона иРНК. Элонгация происходит за счет поочередного присоединения аминокислотных остатков к растущей полипептидной цепи. Терминация наступает, когда синтез полипептида достигает стоп-кодона. У E. coli известны три таких кодона: УАА, УГА и УАГ. В результате действия факторов терминации происходят остановка синтеза белка и диссоциация молекулы иРНК и рибосомы. Скорость синтеза белка в микробной клетке очень велика, так как ДНК бактерий не отграничена мембраной от цитоплазмы, не содержит интронов (участков ДНК, не несущих информации) и соответственно у микробов отсутствуют вырезание их копий из иРНК и сшивание копий экзонов (участков, кодирующих белки). В результате в клетках прокариот не происходит физического разделения процессов синтеза иРНК (транскрипции) и трансляции, поэтому оба процесса часто идут одновременно: трансляция начинается раньше, чем завершена транскрипция. Бактерии также способны одновременно синтезировать несколько идентичных молекул на одной матрице иРНК. При этом иРНК связывается с несколькими рибосомами с образованием комплекса, получившего название «полисомы».
Процесс синтеза белка представляет собой важную мишень для разнообразных антимикробных препаратов. При этом антибиотики имеют различные мишени и механизмы действия, например аминогликозиды и тетрациклины соединяются с малой, а макролиды и линкозамиды - с большой субъединицей рибосом. Белки, синтезируемые клеткой, могут использоваться внутри нее или выделяться в окружающую среду или периплазматическое пространство (грамотрицательные бактерии).
Многие годы считали, что после синтеза на рибосомах молекулы белков сами приобретают нужную форму (третичную структуру). Сейчас мы знаем, что большинство из них приобретают нужную конформацию молекулы с помощью специальных белков, получивших название шаперонов. Молекулы шаперонов не только обеспечивают правильное складывание белков, но и препятствуют неправильному их закручиванию. Эти молекулы абсолютно необходимы для поддержания нормальной жизнедеятельности клетки как эу-, так и прокариот. Процесс укладки белковых молекул является энергетически зависимым и сопровождается расходом энергии АТФ. Количество шаперонов резко возрастает, когда клетка подвергается стрессорному воздействию различных факторов внешней среды (температуры - тепловой шок, токсинов, нарушающих метаболические реакции и др.) При этом шапероны защищают многие белки, включая ДНК-полимеразы от разрушения. Еще одной важной функцией шаперонов является их участие в транспорте белков через мембраны.
Биосинтез нуклеотидов. Пуриновые и пиримидиновые нуклеотиды - это те строительные блоки, из которых синтезируются нуклеиновые кислоты. Кроме того, пуриновые и пиримидиновые нуклеотиды входят в состав многих коферментов и служат для активации и переноса аминокислот, сахаров, липидов в реакциях полимеризации. Исходным соединением для образования пентозной части нуклеотидов служит рибозо-5-фосфат, образующийся в ПФпути. Углеродный скелет пиримидинов происходит из аспартата, который образуется в цикле трикарбоновых кислот. Атомы азота и аминогруппы пуринов и аминосодержащих пиримидинов происходят из аспартата и глутамина.
Ключевым промежуточным продуктом для биосинтеза жирных кислот является ацетилкоэнзим А. Ключевыми промежуточными продуктами для синтеза фосфолипидов является продукт ФДФпути - диоксиацетилфосфат, восстанавливающийся в глицерол-3- фосфат, который соединяется с остатками жирных кислот.
Биосинтез углеводов. Углеводы представлены в бактериальной клетке в виде моно-, ди- и полисахаридов, а также комплексных соединений.
Синтез глюкозы происходит из пирувата за счет обратных реакций, путей распада глюкозы. Для обхода реакций, идущих только в одном направлении, имеются обходные пути, например глиоксилатный цикл.
Дата публикования: 2015-10-09; Прочитано: 6667 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
