- Закон пирамиды энергий, или закон (правило) 10 % Р. Линдемана
- Правило биологического усиления
- Правило «метаболизм и размеры особей», или правило Ю. Одума
- Закон удельной продуктивности
- Правило (принцип) экологического дублирования
- Принцип (правило) эквивалентности В. Тишлера
- Принцип подвижного равновесия А. А. Еленкина
- Принцип продукционной оптимизации Г. Реммерта
- Принцип стабильности
- Правило биоценотической надежности
|
Прежде всего следует напомнить закон (принцип) «энергетической проводимости» (разд. 3.2.3), характерный и для сообществ, и для биоценозов. Иначе не возникла и не сохранилась бы их целостность. Сквозной поток энергии, проходя через трофические уровни биоценоза, постепенно гасится. В 1942 г. Р. Линдеман сформулировал закон пирамиды энергий, или закон (правило) 10%, согласно которому с одного трофического уровня экологической пирамиды переходит на другой, более высокий ее уровень (по «лестнице»: продуцент — консумент — редуцент) в среднем около 10% поступившей на предыдущий уровень экологической пирамиды энергии. Обратный поток, связанный с потреблением веществ и продуцируемой верхним уровнем экологической пирамиды энергии более низкими ее уровнями, например, от животных к растениям, намного слабее — не более 0,5% (даже 0,25%) от общего ее потока, и потому говорить о круговороте энергии в биоценозе не приходится.
Если энергия при переходе на более высокий уровень экологической пирамиды десятикратно теряется, то накопление ряда веществ, в том числе токсичных и радиоактивных, в примерно такой же пропорции увеличивается. Этот факт фиксирован в правиле биологического усиления. Оно справедливо для всех ценозов. В водных биоценозах накопление многих токсичных веществ, в том числе хлорорганических пестицидов, коррелирует с массой жиров (липидов), т. е. явно имеет энергетическую подоснову.
| * Фактически это общесистемый закон: крупные предприятия эффективнее мелких и т. п. Однако в пределах закона оптимальности.
|
При неизменном энергетическом потоке в пищевой сети, или цепи, более мелкие наземные организмы с высоким удельным метаболизмом создают относительно меньшую биомассу, чем крупные*. Значительная часть энергии уходит на поддержание обмена веществ. Это правило «метаболизм и размеры особей», или правило Ю. Одума, обычно не реализуется в водных биоценозах при учете реальных условий обитания в них (в идеальных условиях оно имеет всеобщее значение). Связано это с тем, что мелкие водные организмы в значительной мере поддерживают свой обмен веществ за счет внешней энергии непосредственно окружающей их среды.
На правило Ю. Одума следует обратить пристальное внимание, поскольку из-за антропогенного нарушения природы происходит измельчение «средней» особи живого на суше — крупные звери и птицы истреблены, вообще все крупные представители растительного и животного царства все больше и больше делаются раритетами. Это неминуемо должно вести к общему снижению относительной продуктивности организмов суши и термодинамическому разладу в биосистемах, в том числе сообществ и биоценозов.
Не исключено, что этот разлад служит одним из факторов сбоя в действии принципа Ле Шателье — Брауна (разд. 3.2.3).
Если измельчание особей, согласно правилу Ю. Одума, ведет к производству относительно меньшего объема биомассы, то удельный ее выход с единицы площади в силу более полного заселения пространства увеличивается. Никогда слоны не дадут той биомассы и продукции с единицы площади, какую способна дать саранча и, тем паче, еще более мелкие беспозвоночные. Обсуждаемый эмпирический факт можно назвать законом удельной продуктивности. Кстати, он справедлив для многих системных образований. Так, мелкие предприятия и фермы в сумме могут производить большую хозяйственную продукцию, чем крупные, а тем более крупнейшие, В этом успех мелкого предпринимательства наших дней. Мощные энергоисточники создают тепловые пятна в атмосфере, а потому большие возмущения в ее физике, чем мелкие, равномерно распределенные в пространстве. Именно более полное использование дисперсной энергии составляет подоснову действия закона удельной продуктивности. Этот закон тесно связан с законом оптимальности (разд. 3.2.1). Видимо, его действие сглаживает многие негативные процессы, возникающие в биосфере по вине человека.
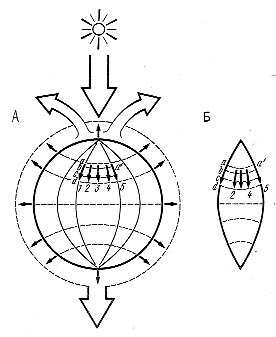 Рис. 3.6. Потоки энергии и механизм обеспечения надежности биотических систем в биосфере: 1, 2, 3... — потоки энергии через виды, а — а1... — связи между ними; А — состояние до уничтожения вида 3; Б — вид 3 исчез, проходившие через него потоки энергии идут через дублирующие виды 2 и 4 Рис. 3.6. Потоки энергии и механизм обеспечения надежности биотических систем в биосфере: 1, 2, 3... — потоки энергии через виды, а — а1... — связи между ними; А — состояние до уничтожения вида 3; Б — вид 3 исчез, проходившие через него потоки энергии идут через дублирующие виды 2 и 4
|
Исчезновение видов, составленных крупными особями, меняет вещественно-энергетическую структуру ценозов. Поскольку энергетический поток, проходящий через биоценоз и экосистему в целом практически не меняется (иначе бы произошла смена типа ценоза), включаются механизмы биоценотического, или экологического, дублирования: организмы одной трофической группы и уровня экологической пирамиды закономерно замещают друг друга. Принципиальная схема такой замены показана на рис. 3.6. Правило (принцип) экологического дублирования можно сформулировать следующим образом: исчезающий или уничтожаемый вид живого в рамках одного уровня экологической пирамиды заменяет другой функционально-ценотический, аналогичный, по схеме: мелкий сменяет крупного, эволюционно ниже организованный более высокоорганизованного, более генетически лабильный и мутабельный менее генетически изменчивого. Поскольку экологическая ниша в биоценозе не может пустовать (разд. 3.8.2), экологическое дублирование происходит обязательно. И действительно, копытных в степи сменяют грызуны, а в ряде случаев растительноядные насекомые. При отсутствии хищников на водоразделах южного Сахалина в бамбучниках их роль выполняет серая крыса. Видимо, таков же механизм возникновения новых инфекционных заболеваний человека. В одних случаях возникает совершенно новая экологическая ниша, а в других борьба с заболеваниями и уничтожение их возбудителей освобождает такую нишу в человеческих популяциях. За 13 лет до открытия ВИЧ (возбудителя СПИДа) была предсказана вероятность появления «гриппоподобного заболевания с высокой летальностью». К сожалению, на докладную записку, поданную мною в Госкомитет по науке и технике никто не обратил внимания, а опубликовать что-то по этому поводу было невозможно из-за жесткой цензуры*.
| * Теоретические основы экосистемного дублирования были сформулированы в статье: Реймерс Н. Ф. Системные основы природопользования//Философские проблемы глобальной экологии. М.: Наука, 1983. С. 121 — 161. Рукопись до опубликования пролежала 10 лет.
|
Экологическое дублирование происходит не только на видовом парном уровне (один вид сменяет другой), но и на уровне сообществ, которые выступают как гамакообразные структуры в технических системах. Такую же роль могут играть пищевые цепи и сети. А поскольку дублирование происходит на основе кибернетического принципа голосования, вместо одного готового блока может включаться другой: сообщество, связанное с одним видом-эдификатором заменяется сообществом, формируемым другим видом-эдификатором. Один вид может быть заменен группой видов (сетевая замена), или наоборот, группа видов исчезает, а вместо них их вещественно-энергетическую функцию станет исполнять всего один вид. Сообщества (синузии, консорции) могут работать как смесители в технических устройствах, где формируется энергетический поток.
Практическое значение экологического дублирования и множественности элементов ценозов очень велико. Известно, что монокультуру, например, гевеи, в тропической зоне вообще невозможно создать из-за неполноты ценозов для ее произрастания (там, где нет ее вредителей, это удается).
Географическое дублирование реализуется согласно принципу (правилу) эквивалентности В. Тишлера (1955): в географически удаленных, но экологически сходных биотопах идентичные биоценотические функции выполняют систематически различные виды, занимающие эквивалентные экологические ниши. В этом легко убедиться, сравнивая конвергентные биоценозы Евразии и Северной Америки. Их называют «изоэкиями» (Г. Гаме, 1918), изоценами или изоценозами (В. Тишлер, 1955). Наличие изоценозов служит одним из важных доказательств энергетического в его основе правила экологического дублирования.
Дублирование — один из механизмов поддержания надежности ценозов. Это наиболее мобильный способ их адаптации. Дублирование может быть и отрицательным — с выпадением видов и части трофических звеньев. Оно может быть полным (очень редко) или частичным, отражающимся лишь на плотности населения или даже лишь характеристиках кроны деревьев, густоты дерновых трав, половозрастном составе популяций животных и тому подобном. Возможны и генетические изменения в популяциях типа усиления хищнических наклонностей у крысы в приведенном для Сахалина примере. Возможно межвидовое и внутривидовое дублирование, а в сельском хозяйстве даже межсортовое. Общий «смысл» остается тем же — максимально полное проведение и использование потока энергии, стабилизация ценоза в меняющихся условиях существования. Это свойство было подмечено А. А. Еленкиным, который в 1921 г. сформулировал принцип подвижного равновесия: биотическое сообщество сохраняется как единое целое вопреки регулярным колебаниям среды его существования, но при воздействии необычных факторов структурно изменяется с переносом «точки опоры» на другие растительные компоненты (группы растений). Если необычные, нерегулярные факторы оказывают многолетнее воздействие, то сообщество формирует иную структуру. Однако следует заметить, что, как правило, сохраняются элементы дублирования в виде малочисленных в ценозе видов, которые могут быть мобилизованы в случае новых резких изменений среды.
Балансовый подход был уточнен сформулированным Г. Реммертом (1978) принципом продукционной оптимизации: отношение между первичной и вторичной продукцией (между продуцентами и консументами) соответствует принципу оптимизации — «рентабельности» биопродукции. Как правило, растения и другие продуценты дают биомассу достаточную, но не излишнюю, для потребления всем биотическим сообществом (с эволюционно определенным запасом, который обеспечивает надежность системы и обычно в 100 раз превышает потребление в экосистемах суши). При относительном «перепроизводстве» органического вещества биоценоз становится «нерентабельным», возникают предпосылки для массового размножения отдельных видов. После периода автоколебаний отношение «популяция — потребление» уравновешивается, биоценоз стабилизируется, балансируются отношения между трофическими уровнями.
Именно благодаря экологическому дублированию, сдвигу в подвижном равновесии и снижению «рентабельности» ценоза возникают массовые размножения нежелательных в хозяйстве организмов. Монокультура в сельском хозяйстве, однопородные и одновозрастные леса с «точки зрения» природы весьма мало рентабельны, неравновесны и поэтому «требуют исправления» массовыми организмами.
Все перечисленные закономерности саморегуляции ценозов обобщаются в виде принципа стабильности: любая относительно замкнутая биосистема с проходящим через нее потоком энергии в ходе саморегуляции развивается в сторону устойчивого состояния. Этот принцип характерен не только для ценозов нижнего уровня иерархии, но и для биосферы в целом. Об этом будет упомянуто в разд. 3.10. Еще раз мы кратко вернемся к принципу стабильности в конце разд. 3.8.3. Тут важно то, что ценоз стремится к нормальной «энергетической проводимости» с помощью механизмов, обобщенно сформулированных в правилах (принципах) экологического дублирования, эквивалентности, подвижного равновесия, продукционной оптимизации и, вероятно, других, еще не открытых исследователями.
Если принцип стабильности справедлив, то парадигма континуума получает еще одно ограничение, а парадигма организма — лишний аргумент своей справедливости. Правда, система может складываться и из ненадежных элементов — см. правило конструктивной эмерджентности (разд. 3.2.1).
Очевидно, возможно сформулировать и обобщающее правило биоценотической надежности: надежность ценоза зависит от его энергетической эффективности в данных условиях среды и возможностей структурно-функциональной перестройки в ответ на изменение внешних воздействий (материала для дублирования, межвидового и внутривидового, поддержания продукционной «рентабельности» и т. п.). Совершенно очевидно, что все эти характеристики ценозов сугубо индивидуальны, но вместе с тем аналогово формируются в сходных условиях среды (принцип эквивалентности). Это дает канву для понимания механизмов функционирования биоценозов, а в случае приложения к одному экологическому биокомпоненту или даже систематической группе, и к сообществу.
Энергетико-продукционные закономерности и способы сохранения экологического равновесия и надежности тесно связаны со структурой биоценозов (как и любых других систем). Попробуем сформулировать имеющиеся обобщения в этой области.

