 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Фотоизомеризация родопсина в фоторецептурной мембране зрительных клеток позвоночных. Проницаемость мембраны и светоиндуцированный электрический сигнал
|
|
Светочувствительные клетки сетчатки – палочки и колбочки генерируют зрительный сигнал в ответ на поглощение квантов света (рис. 8.4). Основной молекулярной фоточувствительной структурой зрительной клетки является фоторецептурная мембрана.

Рис. 8.4. Строение зрительной клетки позвоночных (по L. Stryer, 1987)
В фосфолипидном бислое мембраны встроены молекулы зрительного пигмента родопсина. В темноте ретиналь-хромофор родопсина находится в 11-цис-положении. Поглощение кванта света переводит его в транс-положение
Под действием света резко уменьшается проницаемость мембраны для ионов натрия, один квант закрывает 100-300 каналов. Возникающий светоиндуцированный электрический сигнал гиперполяризации передается к синаптическому окончанию на внутреннем сигменте палочки и приводит к появлению нервного импульса.
Изомеризация ретиналя приводит к закрытию натриевых каналов при участии большого числа молекул медиатора (посредника), находящихся в цитоплазме.
Натриевые каналы способен открывать цГМФ (3, 5 – циклический гуанозин-монофосфат). Между возбуждением родопсина и гидролизом цГМФ фосфодиэстеразой существует сложная многоступенчатая связь.
Фотохимические превращения родопсина. Родопсин – хромопротеин с молекулярной массой порядка 40000. Большой гидрофобный сегмент находится внутри мембраны, гидрофильный сегмент – на поверхности мембраны.
Хромофором является ретиналь, представляющий собой половину молекулы β-каротина. Ковалентная связь ретиналя с опсином осуществляется при образовании альдиминной связи, которая представляет собой протонированное шиффово основание, находящееся в гидрофобном окружении:

В исходном состоянии в родопсине ретиналь находится в 11-цис, 12-в-цис-конфигурации. В результате фотолиза происходит цис-транс-изомеризация хромофора и депротонирование шиффова основания. Эти процессы сопровождаются изменением взаимодействия ретиналя с опсином и конформации последнего. Таким образом, депротонирование шиффова основания служит механизмом его электростатического включения. Основным фактором здесь является нейтрализация заряда боковой цепи Глю 113. Ретиналь, переходя в состояние полностью-транс, запускает этот процесс.
На конечной стадии разрывается альдиминная связь и высвобождается свободный ретиналь в транс-конфигурации. На
Фоторегуляторные и фотодеструкционные процессы
Фоторегуляторные реакции
Действие света на растительные и микробные организмы не ограничивается фотосинтезом, существует большое количество других фотобиологических процессов, среди которых в первую очередь следует выделить процессы фоторегуляции. Особенностью фоторегуляторных систем является их высокая квантовая чувствительность, сочетающаяся с большой эффективностью биологического действия. Это обусловлено триггерным принципом действия систем фоторегуляции, здесь свет служит сигналом, запускающим сложную цепь посмледовательных биохимических изменений, приводящих к определенному фотобиологическому эффекту. Триггерный механизм действия света лежит в основе таких известных фоторегуляторных процессов, как фототропизм, фототаксис, фотоиндуцированный биосинтез пигментов и другие.
Фоторегуляторный процесс включает несколько стадий: поглощение кванта света, образование электронно-воздужденного состояния фоторецептора, фотофизическую реализацию энергии возбуждения, сенсибилизацию фотохимической реакции, образование промежуточных продуктов, конечное проявление фотобиологического эффекта.
Фитохром – наиболее изученный пигмент, участвующий в фоторегуляции растительного организма. С физиологически активной формой фитохрома связаны многообразные изменения в метаболизме, активности ферментов, движениях, росте и дифференциации растений. Считают, что возбужденные красным светом фитохромы через медиаторную систему могут вызвать экспрессию генов, что приводит к синтезу РНК и ферментов.
Фитохром - это хромопротеин, имеет хромофор и белковую часть. Оптические свойства и способность выполнять функцию фоторецептора обусловлены хромофором – линейным тетрапирролом (или билитриеном). Первичная фотохимическая реакция заключается в фотоиндуцированной изомеризации хромофора. Последующие темновые перегруппировки занимают миллисекунды и приводят белковую часть молекулы фитохрома в соответствии с новой конформацией хромофора. Некоторые из индуцированных процессов протекают быстро, другие обладают лагфазой. «Быстрые» процессы обусловлены структурными перестройками мембран, происходят изменения проницаемости для ионов, изменения электрического потенциала и адгезивных свойств поверхности растительных клеток.
В основе механизма «медленных» процессов лежит индуцированное пигментом активирование генов. Изменение проницаемости мембран могут облегчить доступ фитогормонов и других важнейших метаболитов к ДНК. Через определенные медиаторы гормональной природы включаются гены, приводящие к синтезу молекул РНК и ферментов, определяющих направленность той или иной морфогенетической реакции.
К настоящему времени выявлено несколько типов фитохромов, различающихся по структуре белковой части молекулы.
Существуют процессы в проявлении которых наибольшей активностью обладает свет синей области спектра. Например, фототропизм, определяемый как индуцированное односторонним освещением искривление верхушки надземных частей растений. Фототропические изгибы обусловлены различиями в скоростях деления клеток на освещенной и затемненной сторонах растений, что в свою очередь связано с фотоиздуцированным созданием градиента роста – ауксина. Основными рецепторами биологически активного света являются флавиновые хромофоры, способные сенсибилизировать фотоокисление ауксина. Продукты фотолиза ауксина изменяют проницаемость мембран.
Синим светом индуцируются также реакции фоторегуляции движения микроорганизмов, называемые фототаксисом. Фоторецептор локализован в плазматической мембране движущихся водорослей, по-видимому, он представляет собой молекулы типа бактериальных родопсинов. Фотореакция контролирует движение жгутиков.
41. Ультрафиолетовое излучение и фотодеструкционные процессы в ДНК: фотоизомеризация, фотогидротация, образование пиримидиновых аддукатов. Механизмы фотореактивации.
Это инициированные светом реакции, приводящие к таким химическим изменениям молекул, которые нарушают их функции, вызывают повреждения на клеточном и организменном уровнях. Наиболее эффективно индуцируются фотодеструкции коротковолновым ультрафиолетовым излучением (<290 нм). Это излучение хорошо поглощается нуклеиновыми кислотами, белками и другими внутриклеточными компонентами.
Деструктивное действие проявляется в виде реакций фотоокисления, фотоионизации, фотодиссициации и фотоприсоединения.
Например, первичная реакция при УФ-фотолизе триптофана в белках – фотоионизация с образованием катион-радикала АН+ и сольватированного электрона:

В цистиновых остатках – фотодиссициация S-S-связей:

В нуклеиновых кислотах фотоионизация азотистых оснований происходит лишь при поглощении двух квантов УФ-света. Основные фотохимические реакции, в которые вступают основания нуклеиновых кислот: димеризация, гидратация и образование (6-4) пиримидиновых аддукатов (межнитевые ковалентные сшивки).
Деструкционные реакции в большинстве случаев протекают с участием кислорода.
Выделяют 2 типа фотосенсибилизированных деструктивных реакций:
1. Фотовозбужденный сенсибилизатор вступает в окислительно-восстановительные реакции с молекулами, участвуя в переносе электрона или атома водорода. Образуются реакционноспособные радикалы, вступающие в дальнейшие реакции.
2. Происходит перенос энергии от молекулы сенсибилизатора к кислороду с образованием его электронно-возбужденной формы 1О2; образовавшийся синглетный кислород затем окисляет молекулы биологической системы (рис.).

Рис. Фотосенсибилизированные деструктивные реакции с участием кислорода:
где S* — возбужденный фотосенсибилизатор, подвергающийся первичным изменениям, «S+ — окисленная форма сенсибилизатора, А — молекула биосубстрата, участвующая в реакции на последующей стадии.
Фотохимические реакции в ДНК. Считается, что ДНК – основная внутриклеточная мишень при летальном и мутагенном действии УФ-излучения. Волны длиной 260-265 нм совпадают с максимумом в спектре поглощения ДНК.
Основные хромофоры ДНК – это азотистые основания нуклеотидов, причем квантовые переходы пиримидиновых компонентов на порядок выше, чем пуриновых. При поглощении кванта образуются электронно-возбужденные синглетные и триплетные состояния азотистых оснований, энергия которых может превысить энергию ионизации молекулы.
Реакция фотодимеризации ДНК. Считают, что димеры пиримидиновых оснований, образующиеся в одной цепи ДНК при УФ-облучении, - соединяются два основания с образованием кольца циклобутанового типа:

Они составляют 70-80 % от всех летальных повреждений. В ДНК димеризация тимина, по-видимому, идет через синглетное возбужденное состояние основания. Замыкание углеродных связей между двумя молекулами тимина происходит в 5 и 6 положениях, между которыми в соответствии с квантово-механическими расчетами наиболее высока концентрация неспаренных электронов. Это и способствует «разрыву» 5,6-двойной связи и замыканию межтиминовых связей.
Реакция фотогидротации ДНК – фотохимическая реакция пиримидиновых оснований. Происходит под действием УФ-света присоединение воды к пиримидиновому кольцу у С5 и С6 углеродных атомов с разрывом двойных связей, происходит через синглетные возбужденные состояния атомов:
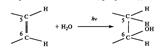
Протекает в одноцепочной ДНК, страдают клетки с активными процессами репликации и транскрипции.
Образование (6-4) пиримидиновых аддукатов – фотопродуктов происходит реже. Могут играть в УФ-мутагенезе важную роль:

Сшивки с белками - межмолекулярные взаимодействия – аминокислотные остатки белков SH- через или ОН-группы прикрепляются к С5 или С6 пиримидиновых оснований ДНК:

Фотореактивация - процесс, направленный на устранение летальных фотопродуктов ДНК при участии фоточувствительного фермента фотолиазы, субстатом которого являются лишь пиримидиновые димеры. Распад димеров пиримидина идет согласно кинетике ферментативных процессов.
Фотолиаза работает в электронно-возбужденном состоянии. Идет перенос электрона от фермента к димеру. При поглощении кванта света фотовозбужденный хромофор фотолиазы донирует электрон димеру с образованием аниона пиримидинового димера. Эта форма не стабильна и распадается на два свободных пиримидина плюс электрон. Восстановление хромофора может идти путем захвата электрона от распавшегося димера.
Фотозащита. Фотоиндуцированное длинноволновым УФ-светом образование в клетке серотонина. Серотонин связывается с ДНК и уменьшает выход димеров, ем самым снижает чувствительность к летальному действию коротковолнового УФ-излучения.
Дата публикования: 2015-11-01; Прочитано: 1256 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
