 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Природа мутаций в области rII
|
|
В модели структуры ДНК Уотсона и Крика предполагается, что замена одной нуклеотидной пары в нормальной нуклеотидной последовательности гена может привести к формированию мутантного фенотипа. Можно предположить, что мутация, в основе которой лежит замена одной нуклеотидной пары, должна обладать следующими свойствами: 1) обратные мутации, переводящие мутантный фенотип в нормальный, должны происходить примерно с той же частотой, что и прямые; 2) ей должна соответствовать определенная точка на генетической карте; 3) такая мутация должна обладать способностью к рекомбинации с любыми другими точечными мутациями, за исключением тех, которые представляют собой независимые замены той же нуклеотидной пары. Некоторые из изученных Бензером rII -мутантов обладали перечисленными свойствами, другие - нет. Данные, представленные в табл. 6.2, показывают, что частота обратных мутаций к дикому типу у различных rII мутантов, способных к рекомбинации друг с другом, сильно различается. Некоторые из rII -мутантов вполне стабильны и не ревертируют к дикому типу (т.е. не дают бляшек на E. coli К (λ)); другие ревертируют к дикому типу с измеримыми и характерными частотами. Генетическая карта rII -мутантов, представленных в табл. 6.2, изображена на рис. 6.3. Результаты рекомбинационного анализа более широкого набора из 60 независимо полученных rII -мутантов представлены на карте
Таблица 6.2. Характеристика восьми rII -мутантов фага Т4
| Мутант | Положение на карте | Частота реверсий |
| r47 | 
| |
| r 104 | 1,3 | 
|
| r101 | 2,3 | 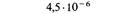
|
| r103 | 2,9 | 
|
| r105 | 3,4 | 
|
| r106 | 4,9 | 
|
| r51 | 6,7 | 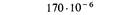
|
| r 102 | 8,3 | 
|
| По Benzer S., 1955. Ргос. Natl. Acad. Sei. USA, 41, 344. |
164 Организация и передача генетического материала
| Рис. 6.3. Генетическая карта rII -мутаций фага Т4, приведенных в табл. 6.2. Цифры над стрелками указывают расстояния. [Benzer S. (1955). Proc. Natl. Acad. Sci., USA, 41, 344.] | 
|

|
| Рис. 6.4. Карта тонкой структуры нескольких участков области rII фага Т4. Числа между стрелками на картах участков а, b, с и d означают процент рекомбинантов дикого типа, возникающих при попарных скрещиваниях между мутантами. [ Benzer S. (1955). Ргос. Natl. Acad. Sci., USA, 41, 344.] |
6. Тонкая структура гена 165
(рис. 6.4). Тщательное изучение этой карты показывает, что некоторые из rII -мутантов (а именно не ревертирующие к дикому типу) рекомбинируют, давая rII + потомство не со всеми остальными rII -мутантами, но лишь с некоторыми из них. Соответственно эти мутации занимают на генетической карте участки большие, чем точечные мутации (размер участка зависит от конкретной мутации), и изображены жирными черными линиями, перекрывающими на карте мутации, с которыми они не рекомбинируют. Например, рассмотрим карту области α на рис. 6.4. Мутант 168 рекомбинирует с мутантами 295 и 312, но ни один из этих трех мутантов не дает рекомбинантов дикого типа с мутантом 47. Все эти четыре мутанта рекомбинируют с мутантами 145, 282 и 228. Если мутанты 168 и 295 - это точечные мутации, то мутант 47 должен возникать в результате изменения более чем одной точки на генетической карте (т.е. изменения более чем одной нуклеотидной пары).
Относительно мутантов типа 47 было показано, что они представляют собой делеции многих последовательных пар нуклеотидов. Исследование рекомбинации двойных rII -мутантов показало, что такие мутации как бы вырезают из генетической карты участки, расположенные между фланкирующими генетическими маркерами, это неудивительно, поскольку у таких мутантов физически отсутствует участок ДНК между этими маркерами (рис. 6.5).
Из этих исследований был сделан важный вывод: фенотипически неразличимые rII -мутации могут быть следствием либо замены отдельной нуклеотидной пары, либо делеции некоторого числа пар нуклеотидов. Свойства, обнаруживаемые у таких делеции, нельзя считать неожиданными. Действительно, вряд ли утрата некоторого числа нуклеотидных пар может быть обратимым процессом, поскольку при этом должны восстанавливаться точное число и последовательность этих пар. Аналогично скрещивание между носителем такой делеции и штаммом с точечной мутацией, расположенной в участке, отсутствующем у партнера по скрещиванию, не должно приводить к появлению рекомбинантов дикого типа. Ни один из геномов, участвующих в таком
| Рис. 6.5. Схема, иллюстрирующая механизм сокращения генетического расстояния между фланкирующими маркерами при скрещивании фаговых мутантов, несущих делецию в одном и том же участке. [ Nomura M., Benzer S. (1961). J. Mol. Biol., 3, 684.] | 
|
166 Организация и передача генетического материала
скрещивании, не содержит нужной нуклеотидной пары в месте точечной мутации. Следовательно, восстановление нуклеотидной последовательности дикого типа невозможно.
Дата публикования: 2014-11-18; Прочитано: 413 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
