 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Глиоксилатный путь катаболизма углеводов. Ферментное обеспечение. Биологическая роль
|
|
ЛИОКСИЛАТНЫЙ ЦИКЛ, циклич. ферментативный процесс, в к-ром происходит превращ. уксусной к-ты в виде ацетилкофермента А (СН3С(О) ~ SKoA) в ди- и трикарбоновые к-ты, а промежут. продуктом является глиоксиловая к-та (см. рис. на стр. 584). Осуществляется в прорастающих семенах масличных растений, а также у нек-рых микроорганизмов (бактерий и плесневых грибов), растущих на среде, которая в качестве единственного источника углерода содержит СН3СООН; у микроорганизмов функционирует наряду с циклом трикарбоновых к-т (ЦТК).
Глиоксилатный цикл локализован в высокоспециализированных субклеточных структурах-глиоксисомах. Образующаяся в них в результате р-ции I глиоксиловая к-та вовлекается снова в цикл, а второй продукт этой р-ции (янтарная к-та) не м. б. использован глиоксисомами и передается в митохондрии, где происходит его окисление до щавелевоукс) сной к-ты. Р-ции глиоксилатного цикла лежат в основе превращения запасного жира в углеводы). В результате 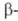 окисления жирных к-т (р-ции II, III) образуется ацетилкофермент А, необходимый для функционирования глиоксилатного цикла.
окисления жирных к-т (р-ции II, III) образуется ацетилкофермент А, необходимый для функционирования глиоксилатного цикла.
При каждом обороте глиоксилатного цикла образуется по одной молекуле янтарной к-ты и НАДН (восстановленная форма кофермента никотинамидадениндинуклеотида). Таким образом, глиоксилатный цикл поставляет для разл. биосинтетич. процессов четырехуглеродные промежут. продукты и энергию, запасенную в НАДН.
Глиоксилатный цикл рассматривают как видоизмененный ЦТК. Последний отличается от глиоксилатного цикла отсутствием р-ций I и IV, к-рые в глиоксилатном цикле катализируются соответствующими ферментами изоцитрат-лиазой и малат-синтазой; у высших растений эти ферменты одновременно присутствуют только в тех тканях, где функционирует глиоксилатный цикл. Остальные р-ции глиоксилатного цикла катализируются теми же ферментами, что и соответствующие р-ции ЦТК.
33. Окислительное фосфорилирование. Система переноса электронов. Участие ферментов. Механизм синтеза АТР при окислительном фосфорилировании – хемиосмотическая теория. АТФ-синтетаза. Регуляция процесса.
Окислительное фосфорилирование – стадия синтеза АТР за счет энергии, запасенной в высокоэнергетических атомах водорода.
Молекулы NADH, NADPH, FADH2, FMNH2, отдают свои высокоэнергетические электроны в дыхательную цепь – в цепь транспорта электронов. Дыхательная цепь состоит из ряда белков с простетическими группами, способными принимать и отдавать электроны. Эти белки располагаются в определенной последовательности и электроны передаются от одного переносчика к другому. При движении высокоэнергетических электронов по дыхательной цепи их энергия затрачивается для синтез АТР. В дыхательной цепи имеются 3 участка, на которых энергия электронов используется для синтеза АТР – эти участки называют пунктами фосфорилирования или пунктами запасания энергии. При движении электронов по дыхательной цепи синтезируется 3 молекулы АТР. В настоящее время общепринятой гипотезой процесса синтеза АТР является хемиосмотическая гипотеза. Первым электрон-транспортным блоком в цепи переноса является NADH-Q-редуктаза, принимающая электроны от NADH и NADPH и передающая их на убихинон (кофермент Q). Убихинон способен также принимать электроны от свободных FADH2, FMNH2. Нужно отметить, что поскольку один из пунктов фосфорилирования находится в участке NADH-Q-редуктазы – электроны от доноров FADH2 и FMNH2 могут обеспечить образование только 2 молекул АТР. Последний блок фосфорилирования в дыхательной цепи называется цитохром-с-оксидазой, которая передает 4е– на О2 с образованием молекулы Н2О.
Дата публикования: 2015-11-01; Прочитано: 1184 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
