 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Общая физиология возбудимых тканей
|
|
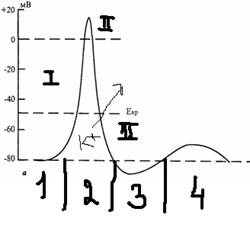
1 фаза – кратковременное повышение возбудимости – в начале развития ПД, когда уже возникла частичная деполяризация, если деполяризация не достигает критического уровня, то регистрируют локальный потенциал.
2 фаза – абсолютная рефрактерная – полная невозбудимость клетки, соответствует пику Пд и продолжается 1-2милисекунду, В этот момент клетка не отвечает на раздражение любой силы, это обусловлено полностью открытыми натриевыми каналами, а те ворота что еще не открыты открываются под действием деполяризации.
3 фаза – относительная рефрактерность – период восстановления возбудимости клетки, когда сильно раздражение может вызвать новое возбуждение. Пониженная возбудимость как следствие все еще повышенной проницаемости для К и избыточным его выходом из клетки, так как выход К препятствует деполяризации клетки.
4 фаза – экзальтация – период повышенной возбудимости. Соответствует следовой деполяризации, Пд можно вызвать достаточно слабым раздражителем.
Скорость протекания фазовых изменений определяет ее лабильность.
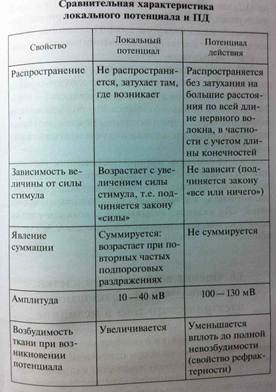
Закон «все или ничего» - по этому закону работает сердечная мышца, подпороговые раздражители не вызывают ответной реакции «ничего», на пороговый раздражитель возникает максимальная ответная реакция «все». Закон неабсолютен, т.к. во-первых, на раздражители подпороговой силы не возникает видимого ответа, но в ткани происходит изменение ПП в виде местного возбуждения. Во-вторых, сердечная мышца, растянутая кровью, при наполнении ею камер сердца реагирует по этому закону, но амплитуда ее сокращения будет увеличиваться по сравнению с сокращением сердечной мышцы, нерастянутой кровью.
4. Критерии для оценки возбудимости тканей: 1. Пороговый потенциал – минимальная величина, на которую надо уменьшить ПП, чтобы вызвать возбуждение в виде ПД. Пороговый потенциал и возбудимость находятся в обратных соотношениях – небольшая величина порогового потенциала свидетельствует о высокой возбудимости клетки.
2. пороговая сила – наименьшая сила раздражителя, способная вызвать возбуждение, при неограниченнии времени ее действия. Реобаза – наименьшая сила электрического тока, способная вызвать импульсное возбуждение при неограниченнии времени ее действия. Если возбудимость ткани высока, реобаза мала. Пороговая сила не зависит от времени ее действия – она зависит только от состояния возбудимости ткани. 3. Пороговое время – это минимальное время в течении которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение, пороговое время также называют полезным временем, чаще используют не его, а хронаксию – наименьшее время, в течении которого должен действовать электрический ток в две реобазы, чтобы вызвать возбуждение.
Лабильность- скорость протекания одного цикла возбуждения, т.е. ПД. Мера лабильности – максимальное число ПД, которое ткань может произвести за 1 секунду. Лабильность ткани понижается при длительном бездействии организма и при утомлении, а также в случае нарушения иннервации. Лабильность нервного волокна 200-40 импульсов в сек., мышечного 100-200, нервно-мышечного 30-50 имп в сек. В эксперименте лабильность исследуют в процессе регистрации максимального числа ПД, которое может воспроизвести клетка при увеличении частоты ритмического раздражения. Чем меньше период рефрактерности, тем выше лабильность и наоборот.
5. Нервное волокно – это отростки нейронов, с помощью которых осуществляется связь нейронов между собой, а также с иннервируемыми клетками и рецепторами. Имеются два типа нервных волокон миелиновые и безмиелиновые. Оболочку безмиелиновых волокон составляют леммоциты(швановские клетки), в которые погружены осевые цилиндры нейронов. Оболочку миелиновых волокон в периферической нервной системе леммоциты, а в ЦНС – олигодендроциты. В миелиновых волокнам мезаксон удлиняется и закручивается вокруг осевого цилиндра, образуя слой миелина вокруг осевого цилиндра. Миелиновая оболочка через равные учатски прерывается, эти перерывы носят название перехваты Ранвье, участки между перехватами – межузловые сегменты. Классификация нервных волокон по Эрлангеру и Гессеру: волокна типа А и В являются миелиновыми, С безмиелиновыми. Волокна А делят на 4 подгруппы – α,β,γ,δ (альфа, бета, гамма, дельта) – это афферентные и эфферентные волокна периферической НС. Волокна типа В преганглионарные волокна ВНС, волокна типа С – постганглионарные волокна ВНС, афферентные волокна от обонятельных, некоторых болевых, тепловых, висцеральных рецепторов. Скорость проведения возбуждения, лабильность, возбудимость понижается от А до С. Для передачи возбуждения на большие расстояния необходимо формирование ПД, проведение возможно лишь в случае наличия на пути проведения потенциалуправляемых ионных каналов, ответственных за формирование новых ПД.
В процессе проведения ПД можно выделить два этапа – этап распространения электрического поля, снижающего мембранный потенциал, и этап генерации новых ПД в новых участках нервного волокна. Непрерывное проведение ПД – только в безмиелиновых волокнах, равномерное распространение ионных каналов по всей длине волокна, центральный участок нервного волокна возбуждается выходящим током от стимулятора, возникает 1 ПД, участок перезаряжается, а соседний участок все еще заряжен -,возникают локальные токи между возбужденным и невозбужденным участками, они вызывают деполяризацию мембраны до Екритического, лавинообразно начинают открываться каналы для Nа, возникают новые ПД в новых участках. Локальные токи распространяются на ограниченные расстояния за счет электрического сопротивления, когда в соседних участках возникнут новые ПД, они вызовут вновь локальный скачок, возникнут новые ПД на соседних участках. Скачкообразное проведение ПД – характерно для миелинизированных волокон – миелин закрывает все ионные каналы на поверхности нервного волокна, кроме перехватов Ранвье, резко увеличено электрическое сопротивление нервного волокна, локальные токи ослабляются значительно меньше и длина скачка возрастает, скорость значительно больше, за счет того что перехваты способны возбуждаться за счет самых слабых локальных токов. Участки между перехватами выполняют роль изолятора, там ПД не возникают. Характеристика проведения возбуждения по нервному волокну:
1. Двустороннее проведение возбуждения – если в эксперименте нанести раздражение в центре нервного волокна, то возбуждение регистрируется как в проксимальном, так и в дистальном отделах.
2. Изолированное проведение возбуждения – изолированное поле ПД каждого волокна гасится межклеточной жидкостью и оболочкой нервных волокон, оно обеспечивает точное афферентное и эфферентное влияние разнородных волокон нерва.
3. Большая скорость проведения возбуждения – достигает 130м/с в Аα, для сравнения в капиллярах 0,5мм/с, эта скорость обеспечивает быстрое влияние на другие нейроны и рабочие органы, а также получение обратной информации.
4. Малая утомляемость нервного волокна – впервые показана Введенским, в его опытах нерв сохранял способность к проведеню возбуждения 6-8 часов беспрерывно, это обусловлено незначительным расходом запасов трансмембранных ионных градиентов, и следовательно нужны небольшие количества АТФ для их восстановления.
5. Возможность функционального блока проведения возбуждения при морфологической целостности волокон – Введенский, при действии различных факторов на нерв, вызывающих длительную деполяризацию, возникает полный блок проведения нервных импульсов(состояние парабиоза) оно связано с инактивацией более 50% натриевых каналов, парабиоз возможен при действии анестетиков, гипоксии, воспалении или охлаждении, после них, если они не вызвали грубых изменений структурных, проведение возбуждения восстанавливается.
6. Высокая лабильность – нервное волокно может проводить 200-400 имп/сек.
6 вопрос. Нервно-мышечный синапс. Возбуждение от нервного волокна к другой клетке
передается с помощью синапса. Нервно-мышечный синапс состоит из пресинаптического окончания, синоптической щели и постсинаптической мембраны. Мотонейроны спинного мозга имеют длинные аксоны, которые в месте контакта с мышцей ветвятся, образуя нервно-мышечные синапсы. Область мышечного волокна, с которой контактирует нервное окончание, носит название концевой пластинки. Пресинаптическое окончание – образовано расширениями по ходу разветвления аксона, иннервирующего мышечное волокно, главный фрагмент – синаптические пузырьки(везикулы), образующиеся в комплексе Гольджи, и доставляющиеся в пресинаптическое окончание, где заполняются медиатором и АТФ, в каждой везикуле от 1 до 10 тысяч молекул химического вещества. Важная структура также митохондрия, осуществляющая энергетическое обеспечение процесса синаптической передачи., цистерны гладкой эндоплазматической сети, содержащие депонированный кальций, микротрубочки и микрофиламенты,участвующие во внутриклеточном движении везикул. Синаптическая щель – содержит межклеточную жидкость и мукуполисахаридное вещество, образующее мостики, обеспечивающие связь пресинаптической и постсинаптической мембран, и может содержать ферменты. Постсинаптическая мембрана – содержит белковые рецепторы, имеющие ионные каналы, и способные связать молекулы медиатора, постсинаптическую мембрану также называют коневой пластинкой. Механизм проведения возбуждения включает 4 этапа. 1 этап – выброс медиатора в синаптическую щель, запускается посредством ПД пресинаптического окончания, деполяризация мембраны пресинаптического окончания ведет к открытию Кальциевых каналов, и он входит согласно электрохимическому градиенту, с помощью кальция происходит выброс ацетилхолина в синаптическую щель. 2 этап – диффузия ацетилхолина к постсинаптической мембране, где находятся N-холинорецепторы, этот процесс занимает 1 миллисекунду. 3 этап – возникновение возбуждения в мышечном волокне. В результате взаимодействия АХ с N-холинорецепторами, открываются ионные каналы и натрий входит в клетку, развивается деполяризация, из-за преобладания натрия, эта деполяризация называется потенциал концевой пластинки, чье электрическое поле достаточно для генерации ПД на мембране близ синапса.
4 этап – удаление Ах из синаптической щели – путем разрушения под действием ацецитхолинэстеразы (расположена в базальной мембране синаптической щели) около 60% захватывается обратно, это делает синтез медиатора более экономичным, часть АХ рассеивается.
Сравнительная характеристика проведения возбуждения по синапсу и нервному волокну:
1. Задержка в передаче сигнала к другой клетке (в нервно-мышечном синапсе 0,5-1 мс) это время расходуется на высвобождение медиатора, диффузию его в постсинаптической мембране, возникновение постсинаптических потенциалов, способных вызвать ПД.
2. Низкая лабильность синапса – в синапсе 30-50 Гц, в нервном волокне 400 Гц, она объясняется тем, что необходимо время на высвобождение медиатора
3. Одностороннее проведение- возбуждение передается только от нервного волокна к другой нервной клетке или к эффекторной клетке, так как пресинаптическое окончание чувствительно только к нервному импульсу, а постсинаптическая мембрана к медиатору
4. Неизолированное проведение – возбуждение рядом расположенных постсинаптических мембран суммируется.
5. Декрементность возбуждения(затухание) – при недостаточном выделении медиатора
6. Проводимость стимулируется и угнетается различными веществами – например кураре и курареподобные вещества связываются с n-холинорецепторами и блокируют действие ах на них.
7. Утомляемость синапсов – ухудшение проводимости вплоть до полной блокады при длительном функционировании синапса, причина истощение медиатора.
7 вопрос Скелетная мышца – произвольная мускулатура, т.к. их движениями можно управлять по собственному желанию. Структурной и сократительной единицей мышцы является мышечное волокно, представляющее собой сильно вытянутое многоядерное волокно, длина его определяется размерном мышцы, имею мембранную оболочку – сарколемму. Внутри мышечного волокна от одного конца до другого тянется масса нитей – миофибрилл, они дают способность к сокращению. Они группируются в пучки по 4-20 штук, в одной миофибрилле насчитывается 2000-2500 тысячи протофибрил – параллельно ледащих нитей двух типов – толстых и тонких, толстые – из миозина, тонкие – из актина. В продольных бороздках нитей миозина расположены регуляторные единицы, состояние из белком тропомиозина и тропонина, эти белки при невозбужденной мышце блокируют контакт актина и миозина, репресорная роль тропонина снимается кальцием, который поступает при возбуждении.
Физиологические свойства мышц: возбудимость(способность возбуждаться под действием раздражителей), проводимость (способность проводить ПД), сократимость (изменение длины или напряжение при возбуждении), растяжимость (спос. Изм. Длину), эластичность (восстановленеи первоначальнйо длины после прекращения ПД). Функции: с помозью движений осущ. Трудовая деятельность, с сокращением мышц связаны не только поддержание позы, но и теплопродукция, кровоток. Классификация двигательных единиц: Представляет собой периферический мотонейрон, его отростки и группу иннервируемых им мышечных волокон. При этом аксон мотонейрона, идущий к мышце, обеспечивающей тонкие движения, иннервируют по 5–12 нервных волокон, тогда как крупные мышцы, осуществляющие массивные движения, получают импульсацию по аксонам, каждый из которых иннервирует 100–500 экстрафузальных мышечных волокон. Их разделяют на медленные (низкопороговые, малые мотонейроны, малоутомляемые) и быстрые(высокопороговые, утомляемые, т.к. неспособны к длительному поддержанию высокочастотного разряда). Мышечные волокна быстрых и медленных также различаются: быстрые волокна белые, более толстые, обладают большей силой, в них меньше митохондрий, жиров и миоглобина, не обладают большой выносливостью. Красные волокна медленные окружены богато капиллярами, получают много крови, тем самым и кислорода. Миоглобин обуславливает цвет волокон, волокна содержат большое количество митохондрий и жиров, это определяет их высокоустойчивость. структурная единица скелетной мышцы – это миофибрилла, а функциональная – мышечное волокно
8 вопрос было обнаружено, что миозиновые нити имею выступы, названные мостиками, они отходят от нитей биполярно, состоят из головки и шейки в спокойном состоянии не могут соединиться с актиновым концом из-за особого расположения тропомиозина, закрывающего активные центры актина. Тропонин подавляет миозин АТФазную активность, что делает невозможным расщепление АТФ, в результате чего мышечные волокна пребывают в расслабленном состоянии. Сокращение мышечных волокон связано с процессами генерации и распространения ПД по поверхностной мембране, а также по мембранам выстилающим трубочки Т, проникая внутрь ПД за счет своего электрического поля приводит к деполяризации мембран продольных трубочек и цистерн саркоплазматического ретикулума, снижение их мембранного потенциала вызывает выход кальция из боковых цистерн в межфибрилярное пространство, и кальций запускает процесс взаимодействия актина с миозином и сокращение мышцы. В присутствии ионов кальция и АТФ тропонин изменяет свою конфигурацию и отодвигает нить тропомиозина, обеспечивая возможность соединения актина и головки миозина. Головка миозина приобретает АТФазную активность, в ней происходит гидролиз АТФ, сопровождаемый пространственной сменой ориентации. Это носит название грибкового движения, обеспечивающее втягивание тонких актиновых миофиламентов в промежутки между толстым миозином, затем отрыв мостика и все заново.
9 вопрос Типы мышечных сокращений. в зависимости от условий, в которых происходит сокращение, различают два типа – изотоническое и изометрическое. Сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным, называется изотоническим. При изометрическом сокращении мышцы укоротиться не может, если оба ее конца укреплены неподвижно, в этом случае напряжение возрастает, а длина мышцы остается неизменной. В естественных условиях сокращения мышц носят смешанный характер. Выделяют одиночное и тетаническое сокращение мышц. Одиночное сокращение изолированной мышцы: оно возникает в эксперименте при раздражении ее одиночным электрическим стимулом. Волна возникает в месте приложения электродов. В изотоническом
режиме одиночное сокращение икроножной мышцы лягушки начинается через короткий скрытый (латентный) период — до 0,01 с, далее следуют фаза подъема (фаза укорочения) — 0,05 с и фаза спада (фаза расслабления) — 0,05—0,06 с. Обычно мышца укорачивается на 5—10 % от исходной длины. Как известно, продолжительность волны возбужения (ПД) мышечных волокон варьирует,
составляя величину порядка 1 — 10 мс (с учетом замедления фазы реполяризации в ее конце). Мышечное волокно реагирует на раздражение по правилу «все или ничего», т.е. отвечает на все надпороговые раздражения стандартным ПД и стандартным одиночным сокращением. Это связано с различной возбудимостью мышечных волокон и разным расстоянием их от раздражающих электродов, что ведет к неодинаковому количеству активированных мышечных волокон. При пороговой силе стимула сокращение мышцы едва заметно, потому что в реакцию вовлекается лишь небольшое количество волокон. При увеличении силы раздражения число возбужденных волокон растет, пока все волокна не окажутся сокращенными, и тогда достигается максимальное сокращение мышцы. Дальнейшее усиление стимулов прироста амплитуды со
кращения не вызывает. Новое сокращение возникает после полного расслабления мышечных волокон. Коэффициентом полезного действия (КПД) называется отношение энергии, затраченной на работу мышц, ко всей энергии, произведенной в мышцах во время работы. АТФ в скелетной
мышце используется для трех процессов: 1) работы натрий-калиевого насоса, обеспечивающего поддержание постоянства градиента концентрации этих ионов по обе стороны мембраны; 2)процесса скольжения акти-новых и миозиновых нитей, приводящих к укорочению миофибрилл (комплекс актин-миозин становится стабильным только при трупном окоченении, когда концентрация АТФ в мышце падает ниже некоторой критической величины); 3) работы кальциевого насоса, активируемого при расслаблении волокна.
10 вопрос Тетаническое сокращение — это длительное слитное сокращение скелетных мышц. В его основе лежит явление суммации одиночных мышечных сокращений. При нанесении на мышечное волокно или целую мышцу двух быстро следующих друг за другом раздражений возникающее сокращение будет иметь большую амплитуду. Сократительные эффекты, вызванные первым и вторым раздражениями, как бы складываются, происходит суммация. При этом в сокращение могут вовлекаться ранее не сокращавшиеся мышечные волокна, если первый стимул вызвал у них подпороговую деполяризацию, а второй увеличивает ее до критической величины. При получении суммации в одиночном волокне важно, чтобы второе раздражение наносилось после исчезновения ПД, т.е. после рефрактерного периода. При сравнительно низких частотах наступает зубчатый тетанус, при большой частоте — гладкий тетанус (рис. 6.4). Их амплитуда больше величины максимального одиночного сокращения. Напряжение, развиваемое мышечными волокнами при гладком тетанусе, обычно в 2—4 раза больше, чем при одиночном сокращении. Режим тетанического сокращения мышечных волокон в отличие от режима одиночных сокращений быстрее вызывает их утомление и поэтому не может поддерживаться длительное время. Из-за укорочения или полного отсутствия фазы расслабления мышечные волокна не успевают восстановить энергетические ресурсы, израсходованные в фазе укорочения. Сокращение мышечных волокон при тетаническом режиме с энергетической точки зрения происходит «в долг». амплитуда гладкого тетануса колеблется в широких пределах в зависимости от частоты стимуляции нерва. При некоторой оптимальной (достаточно высокой) частоте стимуляции амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус получил название оптимума. При дальнейшем повышении частоты стимуляции нерва развивается блок проведения возбуждения в нервно-мышечных синапсах, приводящий к расслаблению мышцы в ходе стимуляции нерва — пессимум Введенского. Частота стимуляции нерва, при которой наблюдается пессимум, получила название пессимальной. В естественных условиях к мышечным волокнам поступают не одиночные, а ряд нервных импульсов, на которые мышца отвечает длительным, тетаническим сокращением, или тетанусом. К тетаническому сокращению способны только скелетные мышцы. При длительном ритмическом раздражении в мышце развивается утомление. Признаками его являются снижение амплитуды сокращений, увеличение их латентных периодов, удлинение фазы расслабления и, наконец, отсутствие сокращений при продолжающемся раздражении, это является следствием того, что калинатриевый насос не успевает возвращать калий и натрий на свои места, натрий накапливается внутри клетки, калий вне клетки, это ведет к деполяризации миоцитов и снижению ПД
11 вопрос Гладкая мышца. Гладкие мышцы построены из веретенообразных одноядерных мышечных клеток. Волокна очень тесно примыкают
друг к другу и связаны между собой низкоом-ными электрическими контактами — нексу
сами. Несмотря на наличие межклеточных щелей шириной 60—150 нм, гладкая мышца функционирует как синцитий — функциональное образование, в котором возбуждение
(медленные волны деполяризации и ПД) способно беспрепятственно передаваться с
одной клетки на другую по крайней мере в пределах одного мышечного пучка, являющегося обычно функциональной единицей гладкой мышцы). Этим свойством гладкая мышца отличается от скелетной и сходна с сердечной, которая тоже представляет собой функциональный синцитий. Однако в сердце достаточно возбудить один миоцит — и возбуждение охватит весь миокард. В гладких мышцах ПД, возникший в одной клетке, распространяется лишь на определенное рассто
яние. ПП в волокнах, не обладающих автоматией, 60—70 мв, она несколько ниже в спонтанно активных клетках — 30-70 мв. ПД - В гладких мышцах внутренних органов регистрируются ПД двух
основных типов: пикоподобные ПД и ПД с выраженным плато. Длительность пикопо-добных ПД составляет 5—80 мс; ПД с плато, характерными для гладких мышц матки, уретры и некоторых сосудов, длятся от 30 до 500 мс. Процесс сокращения гладкомышечных волокон совершается по тому же механизму скольжения нитей актина и миозина относительно друг друга, что и в скелетных мышцах. Того количества кальция, которое входит в клетку при возбуждении, вполне
достаточно для полноценного фазного сокращения. Инициация сокращений гладких мышц с
помощью ионов Са2+ также имеет несколько другой механизм, чем в поперечнополосатых
волокнах. Ионы Са2+ воздействуют на белок кальмодулин, который активирует киназы легких цепей миозина. Это обеспечивает перенос фосфатной группы на миозин и сразу вызывает срабатывание, т.е. сокращение, поперечных мостиков. О существовании тропонин-тропомиозиновой системы сведений не имеется.
12 вопрос Рецептор – структурный элемент клеточной мембраны, воспринимающий действие определнного вида раздражителя и запускающие формирование ответной реакции клетки. По назначению бывают сенсорные, эффекторные и нейронные. сенсорные рецепторы -специфические клетки, настроенные на восприятие различных раздражителей внешней и
внутренней среды организма и обладающие высокой чувствительностью к адекватному
раздражителю. К неадекватным раздражителям — они малочувствительны. Неадекватные раздражители могут возбудить рецепторы: например, механическое давление на глаз вызывает ощущение света, однако энергия неадекватного раздражителя должна быть в миллионы и милли
арды раз больше адекватного. Сенсорные рецепторы являются первым звеном в рефлекторном пути и периферической частью более сложной структуры — анализаторов. эффекторные рецепторы - представляют собой белковые структуры клеточных мембран, а также цитоплазмы и ядра, способные связывать активные химические соединения (гормоны, медиаторы, лекарства и др.) и запускать ответные реакции клетки на эти соединения. Нейронные – постсинаптические рецепторы нейронов, воспринимающие действие медиатора и запускающие процесс обработки информации и ее хранение в нейронах, а также посылку сигналов к другим нейронам ЦНС и приказов (ПД) к рабочим клеткам. Классификация рецепторов в зависимости от вида воспринимающей информации: Механорецепторы возбуждаются при механической их деформации. Они расположены в коже, сосудах, внутренних органах,
опорно-двигательном аппарате, слуховой и вестибулярной системах. Хеморецепторы воспринимают химические изменения внешней и внутренней среды. К ним относятся вкусовые
и обонятельные рецепторы, а также рецепторы, реагирующие на изменение состава крови, лимфы, межклеточной и цереброспинальной жидкости (изменение напряжения 02 и С02, осмолярности, рН, уровня глюкозы и других веществ). Такие рецепторы есть в
слизистой оболочке языка и носа, каротид-ном и аортальном тельцах, гипоталамусе и
продолговатом мозге. Терморецепторы — воспринимают изменения температуры. Они подразделяются на тепловые и холодовые рецепторы и находятся в коже, сосудах, внутренних органах, гипоталамусе, среднем, продолговатом и спинном мозге. Фоторецепторы в сетчатке глаза воспринимают световую (электромагнитную) энергию. Ноцицепторы — их возбуждение сопровождается болевыми ощущениями (болевые рецепторы). Раздражителями этих рецепторов являются механические, термические и химические (гистамин, брадикинин, К+, Н+ и др.) факторы. Болевые стимулы воспринимаются свободными нервными окончаниями, которые имеются в коже, мышцах, внутренних органах, дентине, сосудах. Классификация по расположению: экстеро- и интерорецепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек и органов чувств: зрительные, слуховые, вкусовые, обонятельные, тактильные, кожные болевые и температурные. К интерорецепторам принадлежат рецепторы внутренних органов
(висцерорецепторы), сосудов и ЦНС. Разновидностью интерорецепторов являются рецепторы опорно-двигательного аппарата (проприорецепторы) и вестибулярные рецепторы. В зависимости от степени специфичности рецепторов, т.е. их способности отвечать на один или более видов раздражителей, выделяют мономодальные и полимодальные рецепторы. Мономодальность особенно характерна для экстерорецепторов (зрительных, слуховых, вкусовых и др.) Полимодальные рецепторы приспособлены к восприятию нескольких адекватных раздражителей, например механического и температурного или механического, химического и болевого. К полимодальным рецепторам относятся, в частности, ирритантные рецепторы легких, восприни
мающие как механические (частицы пыли), так и химические (пахучие вещества) раздражители во вдыхаемом воздухе. По структурно-функциональной организации различают первичные и вторичные рецепторы. Первичные представляют собой чувствительные окончания дендрита афферентного нейрона, раздражитель действует непосредственно на окончания сенсорного нейрона. Во вторичных рецепторах имеется специальная клетка, синаптически связанная с
окончанием дендрита сенсорного нейрона, для вторичных рецепторов характерно, что рецепторныи потенциал и потенциал действия возникают в разных клетках, при этом рецепторныи потенциал формируется в специализированной рецепторной клетке, а потенциал
действия — в окончании сенсорного нейрона. Ко вторичным рецепторам относятся слуховые, вестибулярные, вкусовые рецепторы, фоторецепторы сетчатки. По скорости адаптации рецепторы делят на три группы: быстро адаптирующиеся (фазные), медленно адаптирующиеся (тонические)
и смешанные (фазно-тонические), адаптирующиеся со средней скоростью. Свойства рецепторов:
Высокая возбудимость рецепторов. Основное физиологическое значение рецепторов состоит в обеспечении поступления в ЦНС информации о состоянии внешней и внутренней среды, что создает условия для осуществления нервной системой взаимодействия организма и среды. Этому способствует высокая возбудимость рецепторов. Адаптация рецепторов — уменьшение
их возбудимости при длительном действии раздражителя. Процессы адаптации, формирующиеся на разных этапах преобразующей деятельности рецепторов, приводят к снижению амплитуды РП
и как следствие частоты импульсации афферентного нейрона. Спонтанная активность рецепторов. Многие виды рецепторов (фото-, фоно-, вес-тибуло-, термо-, хеморецепторы, проприоре-цепторы) способны генерировать в нейроне импульсацию (потенциалы действия) без
действия на них раздражителя. Эта способность связана со спонтанным колебанием мембранного потенциала в рецепторе, которыи при этом периодически достигает критического уровня деполяризации, что приводит к генерации потенциалов действия в афферентном нейроне.
Возбудимость рецепторов находится под нейрогуморальным контролем целостного организма.
Нервная система может регулировать активность рецепторов через изменение концентрации гормонов, которые, действуя на белки рецепторной мембраны, изменяют состояние ионных каналов и скорость ферментных реакций (например, повышение чувствительности зрительных и слуховых рецепторов под влиянием адреналина, тироксина). Нервная система изменяет чувстви
тельность рецепторов и через регуляцию кровотока в рецепторной зоне, уровень которого
влияет на состав и температуру внеклеточной жидкости около рецептора (например, эндолимфы в органах слуха и равновесия). Вместе с тем нервная система может оказать дорецепторное влияние, регулируя силу действующего на рецептор раздражителя (изменение потока света с помощью зрачкового рефлекса, изменение интенсивности звука с помощью мышц, влияющих на подвижность слуховых косточек и барабанной перепонки).
Педиатрические вопросы
1. Возрастная физиология – это наука, изучающая особенности процесса жизнедеятельности организма на разных этапах онтогенеза. Возрастная физиология тесно связана со многими разделами физиологической науки и, широко использует данные из многих других биологических наук. Так, для понимания закономерностей формирования функций в процессе индивидуального развития человека необходимы данные таких физиологических наук, как физиология клетки, сравнительная и эволюционная физиология, физиология отдельных органов и систем: сердца, печени, почек, крови, дыхания, нервной системы и т. д. Возрастные периоды: Сразу после рождения наступает период, называемый периодом новорожденности (1-10 дней).Основанием для этого выделения служит тот факт, что в это время имеет место вскармливание ребенка молозивом в течение 8-10 дней. грудной период продолжается до года. Начало этого периода связано с переходом к питанию «зрелым» молоком. Во время грудного периода наблюдается наибольшая интенсивность роста, по сравнению со всеми остальными периодами жизни. Период раннего детства длится от 1 года до 4 лет. В конце второго года жизни заканчивается прорезывание зубов. После 2 лет абсолютные и относительные величины годичных приростов размеров тела быстро уменьшаются. С 4 лет начинается период первого детства, который заканчивается в 7 лет. Начиная с 6 лет появляются первые постоянные зубы: первый моляр и медиальный резец на нижней челюсти. Возраст от 1 года до 7 лет называют также периодом нейтрального детства, поскольку мальчики и девочки почти не отличаются друг от друга размерами и формой тела. Период второго детства длится у мальчиков с 8 до 12 лет, у девочек – с 8 до 11 лет. В этот период выявляются половые различия в размерах и форме тела, а также начинается усиленный рост тела в длину. Темпы роста у девочек выше, чем у мальчиков, так как половое созревание у девочек начинается в среднем на два года раньше. Следующий период – подростковый – называется также периодом полового созревания, или пубертатным периодом. Он продолжается у мальчиков с 13 до 16 лет, у девочек – с 12 до 15 лет. В это время наблюдается дальнейшее увеличение скоростей роста – пубертатный скачок, который касается всех размеров тела. Юношеский возраст продолжается у юношей от 18 до 21 года, а у девушек – от 17 до 20 лет. В этот период в основном заканчиваются процесс роста и формирование организма и все основные размерные признаки тела достигают дефинитивной (окончательной) величины. В зрелом возрасте, который продолжается у мужчин от 22 до 60 лет, а у женщин от 21 до 55 лет,форма и строение тела изменяются мало. В пожилом (мужчины – 61-74 года, женщины – 56-74 года) и старческом (75-90 лет) возрасте происходят постепенные инволютивные изменения организма. Выделяют еще один возрастной период – долгожительство (свыше 90 лет).
2,7. Возбудимость мышц плода крайне низка, что обнаруживается при прямом раздражении. У детей также отмечается пониженная возбудимость нервно-мышечной системы — большая пороговая сила и длительная хронаксия. Хронаксия новорожденных в 1,5—10 раз превышает хронаксию мышц взрослых. Развитие возбудимости в онтогенезе непосредственно зависит от становления физико-химических свойств мембран, величины и избирательности их проницаемости для ионов К+, Na+, СГ, т.е. формирования специализированных ионных каналов и ионных насосов. Первым условием возбудимости клеток является наличие потенциала покоя, который составляет для мионов новорожденных 20-40 мВ (у взрослых - 70-80 мВ). У новорожденных отмечается низкий уровень содержания ионов К+ в клетке и большее, чем у взрослых, содержание ионов Na+, поэтому потенциал покоя и потенциал действия значительно меньше, чем у взрослых, более растянут во времени, что определяет большую длительность абсолютной и относительной рефрактерных фаз. В процессе роста (в результате уменьшения проницаемости и улучшения работы ионных помп) потенциал покоя и соответственно потенциал действия возрастают. Низкая лабильность объясняется длительностью абсолютной и относительной рефрактерных фаз, длительностью одиночного мышечного сокращения.
Функции мышц новорожденных детей: основной функцией которой является теплопродукция
3,4. Главный критерий степени зрелости мякотных нервных волокон - их миелинизация, особенно интенсивная к концу антенатального периода. При этом увеличивается расстояние между перехватами Ранвье и изменяется распределение ионных каналов. У ребенка первых лет жизни при неполной миелинизации нервных волокон распределение Na- и К-каналов в мембране равномерное. После завершения миелинизации каналы концентрируются в области перехватов Ранвье, что обусловлено перераспределением в мембране белковых молекул, являющихся основой каналов. В безмякотных волокнах распределение ионных каналов остается равномерным по всей длине волокна. Передние спинномозговые корешки у детей достигают состояния, свойственного взрослым, в период от 2-го до 5-го года жизни, а задние - от 5-го до 9-го года. Миелинизация нервных волокон близка к завершению к 9 годам жизни ребенка.С возрастом в результате созревания нервных волокон их возбудимость, скорость проведения возбуждения и лабильность увеличиваются. Миелинизация нервных волокон обеспечивает увеличение скорости проведения возбуждения вследствие того, что непрерывное проведение возбуждения сменяется сальтаторным и увеличивается расстояние между перехватами Ранвье (ПД электротонически «перепрыгивает» на большее расстояние). Увеличение толщины безмякотного нервного волокна и еще не покрытого миелином мякотного нервного волокна ведет к ускорению проведения возбуждения вследствие уменьшения продольного сопротивления.
5. Структурно-функциональное созревание нервно-мышечных синапсов охватывает пе
риод антенатального и раннего постнаталь-ного периодов развития. Оно имеет свои осо
бенности для пре- и постсинаптических мембран. Созревание пресинаптической мембраны проявляется в увеличении терминального разветвления аксона, усложнении его формы, увеличении площади всего окончания. Степень созревания нервных окончаний значительно увеличиваются к 7—8 годам, при этом проявляются более быстрые и разнообразные движения. В процессе развития усиливается синтез ацетилхолина в мотонейронах, увеличиваются количество активных зон в пресинаптическом окончании и количество квантов медиатора, выделяющегося в синаптическую щель. Когда окончание аксона достигает миотрубки, в соответствующем ее участке появляются скопления митохондрий, рибосом, мик
ротрубок. На поверхности миотрубки образуются выемки, в которых помещается окончание аксона. Образуется примитивное нервно-мышечное соединение. Образуются складки на постсинаптической мембране, в результате чего растет амплитуда ПКП и повышается надежность передачи возбуждения через синапс. Внесинаптическая поверхность мышечного волокна постепенно теряет чувствительность к ацетилхолину. Вследствие незрелости нервно-мышечного синапса у плода и новорожденного синаптическая передача возбуждения происходит медленно. Без трансформации ритма через такой синапс передается не более 20 импульсов в 1 с, а к 7—8 годам жизни — около 100 импульсов в 1 с, т.е. как у взрослого.
6,8. Возбудимость мышц плода крайне низка, что обнаруживается при прямом раздражении. У детей также отмечается пониженная возбудимость нервно-мышечной системы — большая пороговая сила и длительная хронаксия. Хронаксия новорожденных в 1,5—10 раз превышает хронаксию мышц взрослых. Развитие возбудимости в онтогенезе непосредственно зависит от становления физико-химических свойств мембран, величины и избирательности их проницаемости для ионов К+, Na+, СГ, т.е. формирования специализированных ионных каналов и ионных насосов. Первым условием возбудимости клеток является наличие потенциала покоя, который составляет для мионов новорожденных 20-40 мВ (у взрослых - 70-80 мВ). У новорожденных отмечается низкий уровень содержания ионов К+ в клетке и большее, чем у взрослых, содержание ионов Na+, поэтому потенциал покоя и потенциал действия значительно меньше, чем у взрослых, более растянут во времени, что определяет большую длительность абсолютной и относительной рефрактерных фаз. В процессе роста (в результате уменьшения проницаемости и улучшения работы ионных помп) потенциал покоя и соответственно потенциал действия возрастают. Низкая лабильность объясняется длительностью абсолютной и относительной рефрактерных фаз, длительностью одиночного мышечного сокращения. Лабильность. Скелетные мышцы во внутриутробном периоде характеризуются чрезвычайно низкой лабильностью: они способны воспроизводить лишь 3—4 сокращения в секунду. В раннем детском возрасте скелетные мышцы также характери
зуются признаками низкой лабильности. Низкая лабильность объясняется длительностью абсолютной и относительной рефрактерных фаз, длительностью одиночного мышечного сокращения. С возрастом лабильность мышц увеличивается, что ведет, естественно, и к увеличению быстроты движений. Сократимость. Особенностью мышц плода и новорожденных является замедленность одиночных сокращений — как фазы сокращения, так и фазы расслабления, поэтому кривая одиночного мышечного сокращения у новорожденных резко растянута во времени по сравнению с кривой взрослого. Тетанус новорожденного имеет пологое начало и постепенное расслабление, напоминая тетанус утомленной мышцы. Считают, что это связано с более быстрым накоплением метаболитов в мышцах новорожден
ного. Кроме того, у новорожденного, как правило, отсутствуют различия скорости сокращения будущих быстрых и медленных мышц, хотя сами мышцы уже различаются по цвету (белые и красные) и по гистохимическим признакам. Мышцы независимо от частоты и интенсивности раздражителя отвечают тоническим типом сокращения, которое длится столько, сколько продолжается раздражение, без признаков перехода в состояние пессимума. Отсутствие выраженного пессимума связывают с недостаточным структурным оформлении мионевральных синапсов. Эластичность мышц детей раннего возраста значительно больше, чем у взрослых, с возрастом она уменьшается. Упругость и прочность мышц, напротив, с возрастом увеличиваются.
9. Наиболее общим проявлением функции движения является работоспособность мышц, которая лежит в основе возрастной эволюции различных двигательных качеств, опреде-ляющих взаимодействие организма со средой. Напомню, что под физической работо-способностью понимается потенциальная способность человека показать максимум физического усилия в статической, динамической или смешанной работе. Изучение возрастных особенностей величины этого показателя у детей младшего школьного возраста существенно затруднен, так как основной метод регистрации уровня физической работоспособности требует определенного уровня физического развития. Поэтому достоверные данные об изменении мышечной работоспособности относятся почти исключительно к детям старше 6-7 лет. Систематические исследования изменений мышечной работоспособности у детей в возрасте от 7 до 18 лет показывает, что с возрастом работа, выполняемая ребенком на эргографе в течении 1 мин увеличивается, причем прирост количества работы изменяется неравномерно в разные возрастные периоды. Существуют и определенные особенности, характеризующие процесс роста и развития ребенка. Так, например, амплитуде эргограмм свойственно снижение (отчетливое) в период от 7-9 до 10-12 лет, которое сменяется затем постепенным увеличением. Обнаруживается четко выраженное снижение суммарной биоэлектрической активности мышц, то есть с возрастом улучшается использование мышцами нервного напряжения. Изменяется также и характер биоэлектрической активности.Если у детей 7-9 лет пачки импульсов выражены нечетко, часто отмечается непре-кращающаяся электрическая активность, то по мере роста и развития ребенка участки повышенной активности все более разделяются интервалами, на протяжении которых биопотенциалы не регистрируются. Это указывает на то, что с возрастом повышается уровень функционирования двигательного аппарата. По мере роста и развития ребенка происходит концентрация нервных процессов и повышение лабильности мышц.
Одной из важных характеристик мышечной работоспособности является ее восстановление после физической нагрузки.
Дыхательная система.
1. Дыхание — совокупность процессов, обеспечивающих поступление в организм кислорода, использование его для окисления органических веществ с освобождением энергии и выделением углекислого газа в окружающую среду. В среднем в состоянии покоя человек потребляет в течение 1 мин 250 мл 02 и выделяет 230 мл С02. Различают несколько этапов дыхания: 1) газообмен между альвеолами и окружающей средой — вентиляция легких; 2)газообмен между кровью организма и газовой смесью, находящейся в легких; 3)транспорт газов кровью — 02 от легких к тканям, С02 от тканей организма к легким; 4)газообмен между кровью и тканями организма: 02 поступает к тканям, а С02 из тканей в кровь; 5) потребление 02 тканями и выделение С02 — тканевое (внутреннее) дыхание. Совокупность первого и второго этапов дыхания — это внешнее дыхание, обеспечивающее газообмен между окружающей средой и кровью. Оно осуществляется с помощью внешнего звена системы дыхания, включающего легкие с воздухоносными путями и грудную клетку с мышцами, приводящими ее в движение. Прочие этапы дыхания осуществляются посредством внутреннего звена системы дыхания, включающего кровь, сердечно-сосудистую систему, органеллы клеток, и в конечном итоге обеспечивают тканевое (внутреннее) дыхание. Легкие в процессе дыхания выполняют газообменную функцию — главная их роль в организме. Функциональной единицей легкого является ацинус. Кроме газообменной функции, легкие выполняют и ряд других — негазообменных
функций: Выделительная — удаление воды и некоторых летучих веществ: ацетона, этилмер-каптана, этанола, эфира, закиси азота. Газообменная функция является также и выделительной (удаление С02 из организма). Выработка биологически активных веществ гепарина, тромбоксана В2, проста-гландинов, тромбопластина, факторов свертывания крови VII и VIII, гистамина, серотонина, метилтрансферазы, моноаминоксидазы, гликозилтрансферазы. Легкие являются основным источником тромбопластина в организме: когда его мало в крови, выработка возрастает, когда много — выработка тром
бопластина уменьшается. Инактивация биологически активных веществ — эндотелий капилляров легких инак-тивирует за счет поглощения или ферментативного расщепления многие биологически активные вещества, циркулирующие в крови. Защитная функция — легкие являются барьером между внутренней и внешней средой организма, в них образуются антитела, осуществляется фагоцитоз, вырабатываются лизоцим, интерферон, лактоферрин, иммуноглобулины; в капиллярах задерживаются и разрушаются микробы, агрегаты жировых клеток, тромбоэмболы. Функцию фагоцитоза выполняют так называемые альвеолярные фагоциты. Это крупные клетки округлой формы, они находятся в свободном виде в альвеолах и являются макрофагами. Эти клетки фагоцитируют попавшие в легкие мелкие пылевые частицы (менее 2 мкм), микроорганизмы и вирусы, компоненты сур-фактанта, продукты распада клеток альвеолярного эпителия. Макрофаги с воздухом достигают бронхиол, а далее с помощью мерцательного эпителия с мокротой выделяются из организма или попадают в желудочно-кишечный тракт. Терморегуляция — в легких вырабатывается большое количество тепла. Легкие являются резервуаром воздуха для голосообразования.
Воздухоносный путь — это пространство, которое обеспечивает доставку атмосферного воздуха в газообменную область. Негазообменные функции: Очищение вдыхаемого воздуха от крупных пылевых частиц происходит в волосяном фильтре в преддверии носа. В полости носа удаляется до 85 % мелких частиц (размер до 4,5 мкм). Более мелкие частицы (до 1 мкм), прошедшие дальше, оседают на слизистой оболочке носа, чему способствуют турбулентный характер движения воздуха в носовых ходах и слизистый секрет оболочки носа. Очищению воздуха от попавших частиц способствуют кашель и чиханье — защитные рефлексы. Кашлевой рефлекс начинается с глубокого вдоха.
Он возникает в результате раздражения ирритантных рецепторов слизистой оболочки
гортани, глотки, трахеи и бронхов, импульсы от которых поступают по верхнегортанному,
тройничному и блуждающему нервам в ядро солитарного тракта, а оттуда — к экспиратор
ным нейронам, обеспечивающим форсированный выдох через рот и выброс воздуха с
попавшими в воздухоносные пути частицами. Главную роль в осуществлении кашлевого рефлекса играют мышцы живота. Чиханье возникает в результате раздражения ре
цепторов тройничного нерва оболочки носа. Механизм чиханья подобен механизму
кашля, но воздух изгоняется в основном через нос. При этом возникает слезоотделение, что способствует очищению воздуха: попавшие в нос частицы смываются слезой, оттекающей через слезноносовой канал в полость носа. Увлажнение вдыхаемого воздуха достигает 100 %, начинается еще в верхних дыхательных путях, в первую очередь в полости носа насыщением воздуха влагой слизистой оболочки. Слизь образуется в результате фильтрации жидкости из кровеносных капилляров, выделений из желез слизистой оболочки и слезных желез. За сутки из оболочки носа может испаряться в зависимости от температуры и влажности воздуха до 0,5 л воды. Согревание воздуха также начинается в верхних дыхательных путях, в альвеолы воздух поступает при температуре 37 °С. Особое значение в согревании вдыхаемого воздуха имеет слизистая оболочка носа, богато снабженная кровеносными сосудами. При дыхании носом температура воздуха уже в носоглотке доходит до 35—36 °С. При раздражении чувствительных окончаний тройничного нерва холодным воздухом афферентные импульсы поступают к парасимпатическим центрам продолговатого мозга, в результате
чего расширяются сосуды оболочки носа и воздух лучше нагревается. Этому способствует также сужение носовых ходов в результате увеличения объема кавернозной ткани носовых раковин: воздух проходит более тонкой струей и лучше согревается. Если температура воздуха выше 37 °С, то он охлаждается до этой температуры. Таким образом, нос как начальный отдел воздухоносных путей играет главную роль в очищении, согревании и увлажнении вдыхаемого воздуха. Воздухоносные пути участвуют в процессах терморегуляции за счет теплоиспарения, конвекции и теплопродукции. Грудная клетка является герметической полостью для легких. Она предохраняет от высыхания и механического повреждения. Грудная клетка своими экскурсиями обеспечивает сужение и расширение легких, а значит, их вентиляцию.
1. Важную роль в процессах внешнего дыхания играет отрицательное давление в плевральной щели. Плевральная щель — узкое пространство между висцеральной и париетальной листками плевры, заполненное серозной жидкостью, обеспечивающей прилегание двух листков плевры друг к другу и скольжение их относительно друг друга. Отрицательное давление — это величина, на которую давление в плевральной щели ниже атмосферного. В норме это (—4)—(—8) мм рт.ст. Таким образом, реальное давление в плевральной щели составляет величину порядка 752—756 мм рт.ст. и зависит от фазы дыхательного цикла. При максимальном вдохе отрицательное давление возрастает до —20 мм рт.ст., при максимальном выдохе оно приближается к нулю (особенно в нижних отделах), т.е. становится почти равным атмосферному давлению (760 мм рт.ст.). Рост легких в процессе развития организма отстает от роста грудной клетки. Поскольку на легкое атмосферный воздух действует только с одной стороны — через воздухоносные пути, оно растянуто и прижато к внутренней стороне грудной клетки. Вследствие растянутого состояния легких возникает сила, стремящаяся вызвать спадение легких. Эта сила называется эластической тягой легких (ЭТЛ) Так как плевральная щель не сообщается с атмосферой, давление в ней ниже атмосферного на величину ЭТЛ: при спокойном вдохе — на 8 мм рт.ст., при спокойном выдохе — на 4 мм рт.ст. О том, что легкие находятся в растянутом состоянии, свидетельствует факт спадения их при пневмотораксе (патологическом состоянии, возникающем при нарушении герметичности плевральной щели, в результате чего атмосферный воздух заполняет плевральную щель, оказываясь между висцеральным и париетальным листками плевры; ЭТЛ формируют эластиновые и коллаге-новые волокна, гладкие мышцы сосудов легких и, главное, поверхностное натяжение пленки жидкости, покрывающей внутреннюю поверхность альвеол. Силы поверхностного натяжения составляют 2/3 величины ЭТЛ. Величина поверхностного натяжения альвеолярной пленки существенно изменяется в присутствии сурфактанта — активного вещества легких, образующего слой толщиной 50 нм внутри альвеол, альвеолярных ходов, мешочков и бронхиол. Сурфактант содержит фосфолипиды (в частности, лецитин), триглицериды, холестерин, протеины и углеводы. Роль: Уменьшает поверхностное натяжение жидкости, покрывающей альвеолы, примерно в 10 раз, тем самым предотвращая ателектаз (слипание) альвеол. Выполняет защитную роль: а) обладает бактериостатической активностью; б) обеспечивает обратный транспорт пыли и микробов по воздухоносному пути; в) защищает стенки альвеол от повреждающего действия окислителей и перекисей; г) уменьшает проницаемость легочной мембраны, что является профилактикой развития отека легких — это достигается уменьшением выпотевания жидкости из крови в альвеолы. У курильщиков защитные свойства сурфактанта ослабевают, уменьшается активность альвеолярных макрофагов, снижаются защитные функции легких в целом, чаще встречаются заболевания легких и других органов. Облегчает диффузию кислорода из альвеол в кровь вследствие хорошей растворимости кислорода в нем. Значение отрицательного давления в плевральной щели для организма заключается в том, что оно обеспечивает сжатие грудной клетки при выдохе и куполообразное положение диафрагмы, так как давление в брюшной полости несколько выше атмосферного за счет тонуса мышц стенки живота, а в грудной полости оно ниже атмосферного.
2. Механизм вдоха и выхода. Поступление воздуха в легкие при вдохе и изгнание его из легких при выдохе осуществляются благодаря ритмичному расширению и сужению грудной клетки. Вдох является первично активным (осуществляется с непосредственной затратой энергии), выдох также может быть первично активным, например, при форсированном дыхании. При спокойном же дыхании выдох является вторично активным, так как осуществляется за счет потенциальной энергии, накопленной при
вдохе. Три одновременно протекающих процессов: 1) расширение грудной клетки, 2)расширение легких, 3) поступление воздуха в альвеолы. Расширение грудной клетки при вдохе обеспечивается сокращением инспираторных мышц и происходит в трех направлениях: вертикальном, фронтальном и сагиттальном. Инспираторными мышцами являются диафрагма, наружные межреберные и межхрящевые. В вертикальном направлении грудная клетка расширяется в основном за счет сокращения диафрагмы и смещения ее сухожильного центра вниз. Диафрагмальная мышца — главная дыхательная мышца, в норме вентиляция легких на 2/3 осуществляется за счет ее движений. Диафрагма принимает участие в обеспечении кашлевой реакции, рвоты, икоты, в родовых схватках. При спокойном вдохе купол диафрагмы опускается примерно на 2 см, при глубоком вдохе — до 10 см. Расширение грудной клетки в переднезаднем направлении и в стороны происходит при поднятии ребер вследствие сокращения наружных межреберных и межхрящевых мышц. Наружные межреберные мышцы при своем сокращении с одинаковой силой тянут верхнее ребро вниз, а нижнее поднимают вверх. энергия при вдохе расходуется только на частичное преодоление ЭТЛ и брюшной стенки, а ребра поднимаются сами. При этом расширяющаяся грудная клетка способствует также преодолению ЭТЛ. При расширении грудной клетки движение нижних ребер оказывает большее влияние на ее объем и вместе движением диафрагмы вниз, обеспечивает лучшую вентиляцию нижних долей, чем верхушек легких. Вместе с расширением грудной клетки расширяются и легкие. Главная причина расширения легких при вдохе — атмосферное давление воздуха, действующее на легкое только с одной стороны; вспомогательную роль выполняют силы сцепления (адгезии) висцерального и париетального листков плевры. Увеличение отрицательного давления в плевральной щели является не причиной, а следствием расширения легких.
Имеется еще одна сила, которая способствует расширению легких при вдохе, — это
сила сцепления между висцеральным и париетальным листками плевры. Но она крайне мала по сравнению с атмосферным давлением. Поступление воздуха в легкие при их
расширении является результатом некоторого (на 1,5 мм рт.ст.) падения давления в альвеолах. Этого градиента давления оказывается достаточно, поскольку воздухоносные
пути имеют большой просвет и не оказывают существенного сопротивления движению
воздуха. Кроме того, увеличение ЭТЛ при вдохе обеспечивает дополнительное расши
рение бронхов. Вслед за вдохом плавно начинается выдох. (Экспираторными мышцами являются внутренние межреберные мышцы и мышцы брюшной стенки) Спокойный выдох осуществляется без непосредственной затраты энергии. При вдохе растягиваются легкие, вследствие чего возрастает ЭТЛ. Как только прекращается поступление
нервных импульсов к мышцам вдоха по диафрагмальному и межреберным нервам, прекращается возбуждение мышц, вследствие чего они расслабляются. Грудная клетка суживается под влиянием ЭТЛ и постоянно имеющегося тонуса мышц стенки живота
при этом органы брюшной полости оказывают давление на диафрагму. Вследствие сужения грудной клетки легкие сжимаются. Поднятию купола диафрагмы способствует
также ЭТЛ. Давление воздуха в легких возрастает на 1,5 мм рт.ст. в результате уменьшения их объема, воздух из легких изгоняется в атмосферу. Сужению грудной клетки (опусканию ребер) способствует ее масса. Но главную роль играет ЭТЛ, настолько сильно сжимающая грудную клетку при выдохе, что при вдохе она расправляется сама, без непосредственной затраты энергии за счет сил упругости (потенциальной энергии), накопленной при выдохе.
3. Вентиляция легких – газообмен между атмосферным воздухом и легкими. Гипервентиляция – произвольное усиление дыхания, не связанное с метаболитическими потребностями организма. Гиперпное – непроизвольное усиление дыхания с связи с реальными потребностями организма. Дыхательный объем (ДО) — это объем воздуха, который человек вдыхает и выдыхает при спокойном дыхании, при этом продолжительность одного цикла дыхания составляет 4—6 с, акт вдоха проходит несколько быстрее. Такое дыхание называется эйпное (хорошее дыхание). Резервный объем вдоха (РО вдоха) — максимальный объем воздуха, который человек может дополнительно вдохнуть после спокойного вдоха. Резервный объем выдоха (РО выдоха) — максимальный объем воздуха, который можно выдохнуть после спокойного выдоха. Остаточный объем (00) — объем воздуха, остающийся в легких после максимального выдоха. Жизненная емкость легких (ЖЕЛ) — это наибольший объем воздуха, который можно выдохнуть после максимального вдоха. У молодых людей должную величину ЖЕЛ можно рассчитать по формуле: ЖЕЛ = Рост (м) • 2,5 [л]
Функциональная остаточная емкость (ФОЕ) — количество воздуха, остающееся в
легких после спокойного выдоха, равна сумме остаточного объема и резервного объема выдоха. Общая емкость легких (ОЕЛ) — объем воздуха, содержащийся в легких на высоте максимального вдоха, равна сумме ЖЕЛ плюс остаточный объем. Общая емкость легких, как и другие объемы и емкости, весьма вариабельна и зависит от пола, возраста и роста. Так, у молодых людей в возрасте 20—30 лет она равна в среднем около 6 л, у мужчин в 50—60 лет — в среднем около 5,5 л. Минутный объем воздуха (MOB) — это объем воздуха, проходящего через легкие за 1 мин. Он составляет в покое 6—8 л, частота дыхания — 14—18 в 1 мин. При интенсивной мышечной нагрузке MOB может достигать 100 л. Максимальная вентиляция
легких (МВЛ) — это объем воздуха, который проходит через легкие за определенный промежуток времени при максимально возможной глубине и частоте дыхания. МВЛ
может достигать у молодого человека 120—150 л/мин, а у спортсменов — 180 л/мин, он
зависит от возраста, роста, пола. При прочих равных условиях МВЛ характеризует проходимость дыхательных путей, а также упругость грудной клетки и растяжимость
легких. Газообмен в легком, естественно, ведет к изменению газового состава в легком
по сравнению с составом атмосферного воздуха. В покое человек потребляет около
250 мл 02 и выделяет около 230 мл С02. Поэтому в альвеолярном воздухе уменьшается количество 02 и увеличивается — С02. Изменения содержания 02 и С02 в альвеолярной смеси газов являются следствием потребления организмом 02 и выделения С02.
В выдыхаемом воздухе количество 02 несколько возрастает, а С02 уменьшается по
сравнению с альвеолярной газовой смесью вследствие того, что к ней добавляется воздух воздухоносного пути, не участвующий в газообмене и, естественно, содержащий С02 и 02 в таких же количествах, как и атмосферный воздух. Азот в газообмене не участвует, некоторое увеличение содержания его в альвеолярном воздухе является относительным: объем выдыхаемого воздуха несколько меньше объема вдыхаемого. Это объясняется тем, что С02 выделяется из организма несколько меньше, нежели потребляется 02, из-за различного содержания углерода и кислорода в различных окисляемых вещест
Дата публикования: 2014-11-19; Прочитано: 5205 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
