 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Кафедра Зоологии
|
|
-----------------------------------------------------------------------------
Федеральное государственное бюджетное образовательное учреждение
Высшего профессионального образования
Ульяновский государственный педагогический университет
имени И. Н. Ульянова
Естественно-географический факультет
Кафедра Зоологии
Отчет
«Полевая практика по популяционной генетике»
Работу выполнили:
студенты 4 курса
естественно-географического
факультета
гр. БЭ-08-1
Антошин Д., Шакин М.
Проверил:
Мищенко А.В.
г. Ульяновск 2012
Содержание:
1.изучение закономерностей модификационной изменчивости.
2.Изучение мутационной изменчивости.
3.цитоплазматическое наследование.
4.Закон гомологических рядов.
5.Центры происхождения растений.
6.Изучение изменчивости седых пятен у клевера.
7.Изучение изменчивости в популяциях колорадского жука.
1.Изучение закономерностей модификационной изменчивости.
Модификационная изменчивость - это эволюционно закрепленные реакции организма на изменения условий внешней среды при неизменном генотипе. Такой тип изменчивости имеет две главные особенности. Во- первых, изменения затрагивают большинство или все особи в популяции и у всех них проявляются одинаково. Во-вторых, эти изменения обычно имеют приспособительный характер. Как правило, модификационные изменения не передаются следующему поколению. Классический пример модификационной изменчивости дает растение стрелолист, у которого надводные листья приобретают стреловидную форму, а подводные - лентовидную.
Если у гималайского кролика на спине удалить белую шерсть и поместить его в холод, на этом месте вырастет черная шерсть. Если черную шерсть удалить и наложить теплую повязку, вырастет белая шерсть. При выращивании гималайского кролика при температуре 30*С вся шерсть у него будет белая. У потомства двух таких белых кроликов, выращенного в нормальных условиях, появится "гималайская", окраска. Такая изменчивость признаков, вызванная действием внешней среды и не передающаяся по наследству, называется модификационной.
Обычно, говоря о модификационных изменениях, имеют в виду морфологические изменения (например, изменение формы листьев) или изменения окраски). Однако нередко в эту группу включают и физиологические реакции. Регуляция работы генов лактозного оперона кишечной палочки представляет собой пример такой физиологической реакции. Напомним, в чем она состоит. При отсутствии в среде обитания бактерий глюкозы и при наличии лактозы бактерия начинает синтезировать ферменты для переработки этого сахара. Если же в среде появляется глюкоза, эти ферменты исчезают и бактерия возвращается к стандартному обмену веществ.
Другой пример физиологической реакции - увеличение числа эритроцитов в крови у человека, поднявшегося в горы. Когда человек спускается вниз, где содержание кислорода нормально, число эритроцитов возвращается к норме.
В обоих примерах модификационные изменения имеют ясно выраженный приспособительный характер, поэтому их часто называют физиологическими адаптациями.
Модификационная изменчивость не затрагивает наследственной основы организма - генотип и поэтому не передается от родителей потомству.
Еще одна особенность модификационной изменчивости - ее групповой характер. Определенный фактор внешней среды вызывает сходное изменение признаков у всех особей данного вида, породы или сорта: под воздействием ультрафиолетовых лучей все люди загорают, все растения белокочанной капусты в жарких странах не образуют кочана. При этом, в отличие от мутаций, модификации направленны, имеют приспособительное значение, происходят закономерно, их можно предсказать. Если листья на деревьях уже распустились, а ночью были заморозки, то утром листья у деревьев примут красноватый оттенок. Если мышей, которые жили на равнинах вблизи гор, переселить в горы, то у них повысится содержание гемоглобина в крови.
Благодаря возникновению модификаций особи непосредственно (адекватно) реагируют на изменение условий среды и лучше приспосабливаются к ней, что дает возможность выжить и оставить потомство.
Входе проведения исследований было произведено измерение длины листовой пластинки (Х) и количества простых листьев (Z) шиповника белого(Rosa alba).
Результаты были занесены в таблицу вариантов.
| Х Z | ||||||||||
X-длина листовой пластики (мм)
Z-количество простых листьев
1.Определяем границу вариационного ряда, находим min и max значения.
lim (X) max=134
lim (X) min=22
lim (Z) max=11
lim (Z) min=5
2. Определяем размах изменчивости.
(X) max-min=134-22=112
(Z) max-min=11-5=6
3. Определяем классовый промежуток.
max-min/K=L
(X) 112/10=11.2=11
(Z) 6/10=0.6=1
4. Построить вариационный ряд
| классы | n | |||||||||||
| среднее | ||||||||||||
| X | 22-32 | 33-44 | 44-54 | 55-65 | 66-76 | 77-87 | 88-98 | 99-109 | 110-120 | 121-131 | 132-142 | |
| Y(x) | ||||||||||||
| отклонение | -2 | -1 | Xo |
| классы | n | |||||||
| среднее | ||||||||
| Y(Z) | ||||||||
| отклонение | -2 | -1 | Zo |
5.Строим вариационную кривую
(X)
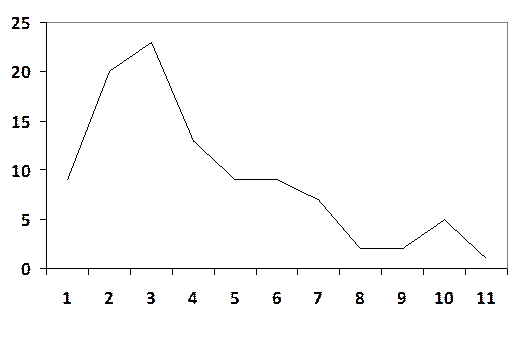

(Z)
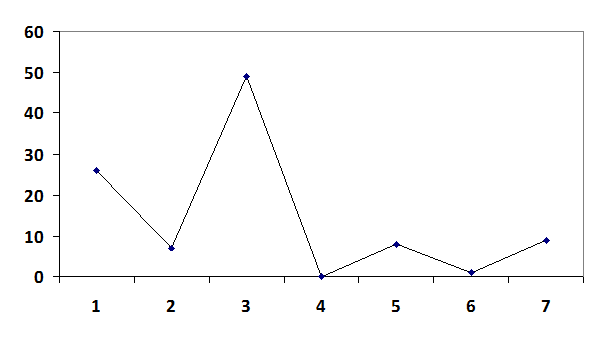
6. Находим среднее арифметическое выборки.
X= Xo+(Σ(X-Xo)/n)*l
| (X) | ||||||||
| + | ||||||||
| - | ||||||||
| + | ||||||||
| - |
Σ(X-Xo)=113
X(в)=49+113/100*11=49+12.43=61.43
Проверка:
| (X) | |||||||
| + | |||||||
| - | |||||||
| + | |||||||
| - |
Σ(X-Xo)=13
Х(в)=60+13/100*11=60+1.43=61.43
Z= Zo+(Σ(Z-Zo)/n)*l
| (Z) | ||||
| + | ||||
| - | ||||
| + | ||||
| - |
Σ(Z-Zo)=-4
Z(в)=7-4/100*1=7-0.04=6.96
Проверка:
| (Z) | |||
| + | |||
| - | |||
| + | |||
| - |
Σ(Z-Zo)=-104
Z(в)=7-104/100*1=8-1.04=6.96
7.Находим величину стандартного отклонения,среднеквадратичного отклонения (Ϭ).
(X)
Ϭ=±√(Σ(X-Xo)²/n)*l
Ϭ=±√((Σ(X-Xo)²-[ Σ(X-Xo)]²/n)/n)*l
| (X) | 1² | 2² | 3² | 4² | 5² | 6² | 7² | 8² |
Σ(X-Xo)²=729
Ϭ(X)= ±√(729-113²/100)/100*11=±√(729-127.69)/100*11=±√601.31/100*11=
±√6.0131*11=2.45*11=26.95
Проверка:
| (X) | 1² | 2² | 3² | 4² | 5² | 6² | 7² |
Σ(X-Xo)²=603
Ϭ(X)= ±√(603-13²/100)/100*11=±√(603-169)/100*11=±√601.31/100*11=
±√6.0131*11=2.45*11=26.95
(Z)
Ϭ=±√(Σ(Z-Zo)²/n)*l
Ϭ=±√((Σ(Z-Zo)²-[ Σ(Z-Zo)]²/n)/n)*l
| (Z) | 1² | 2² | 3² | 4² |
Σ(Z-Zo)²=296
Ϭ(Z)= ±√(296-(-4)²/100)/100*1=±√(296-0.16)/100*1=±√295.85/100*1=1.72
Проверка:
| (Z) | 1² | 2² | 3² |
Σ(Z-Zo)²=404
Ϭ(Z)= ±√(404-104²/100)/100*1=±√(404-108.16)/100*1=±√295.85/100*1=1.72
8. Находи ошибку среднего (m)
(X)
m=Ϭ/-√n
Хп=Хв±2*m
m(X)=26.95/-√100=-2.695 Х=61.43+2*(-2.695)=56.04
Х=61.43-2*(-2.695)=66.82
(Z)
m=Ϭ/-√n
Zп=Zв±2*m
m(Z)=1.72/-√100=-0.172 Z=-4+2*(-0.172)=-4.344
Z=-4-2*(-0.172)=-3.656
9. Находим коэффициент вариации, размах изменчивости признака для популяции.
(X)
C.V=Ϭ/x*100%
C.V(X)=Ϭ/x*100%=26.95/61.43*100%= 43,87
X±3Ϭ=61,43±80,85=142,28
=-19,42
(Z)
C.V=Ϭ/z*100%
C.V(Z)=Ϭ/z*100%=1.72/6.96*100%= 24,71
Z±3Ϭ=6,96±5,16=12,12
=1,8
Выводы:
(Х)
- Среднее значение признака длины листовой пластинки (Х) в выборке равно 61,43 мм.
- Среднее значение листовой пластинки (Х) в пределах популяции 61,43±2*2,695мм
- Изменчивость признака (Ϭ) равна 26,95мм
- Коэффициент вариации по длине листовой пластинки равен 43,87
- Среднее значение изменчивости длины листовой пластинки колеблется 61,43±80,85
(Z)
- Среднее значение признака числа простых листьев (Z) в выборке равно -4 мм.
- Среднее значение листовой пластинки (Z) в пределах популяции -4±2*0,172мм
- Изменчивость признака (Ϭ) равна 1,72мм
- Коэффициент вариации по числу простых листьев равен 24,71
- Среднее значение изменчивости числа простых листьев колеблется -4±5,16
Корреляционный лист.
| X Z | Y(Z) | Z-Zo | Σ(X-Xo) | Σ(Z-Zo)*(X-Xo) | ||||||||||||
| 22-32 | 33-43 | 44-54 | 55-65 | 66-76 | 77-87 | 88-98 | 99-109 | 110-120 | 121-131 | 132-142 | ||||||
| -1 | -4 | |||||||||||||||
| -2 | -19 | |||||||||||||||
| Y(X) | ||||||||||||||||
| X-Xo | -2 | -1 | ||||||||||||||
| Σ(Z-Zo) | -10 | -20 | -25 | -3 | ||||||||||||
| Σ(X-Xo)* *(Z-Zo) | -3 |
R=(Σ(X-Xo)*(Z-Zo)- Σ(X-Xo)*(Z-Zo)/n)/√ Σ(X-X)²*(Z-Z)²=(339-113*(-4)/100)/√(729-127,69)*(296-0,16)=343,52/√177900,42=343,52/421,78=0,8
Вывод: Коэффициент r=0,8 показывает коррелятивную устойчивость зависимости признаков.
2.Изучение мутационной изменчивости
Термин "мутация" восходит к латинскому слову "mutatio", что в буквальном переводе означает - изменения или перемена. Мутационная изменчивость обозначает устойчивые и явные изменения генетического материала, что выводится в наследственные признаки. Именно это является первым звеном в цепочке формирования наследственных болезней и патогенеза. Данное явление стало активно изучаться только во второй половине 20-го века, а в настоящее время всё чаще можно слышать, что мутационная изменчивость должна изучаться, так как знание и понимание данного механизма становится ключевым для преодоления проблем человечества.
Существует несколько видов мутаций в клетках. Их классификация зависит от разновидности самих клеток. Генеративные мутации происходят в половых клетках, также существуют гаметические клетки. Любые изменения наследуются и зачастую обнаруживаются в клетках потомков, от поколения к поколению передаётся ряд отклонений, которые в итоге становятся причиной заболеваний.
Соматические мутации относятся к неполовым клеткам. Их особенность в том, что они проявляются только у того индивида, у которого появились. Т.е. изменения не передаются по наследству другим клеткам, а только при делении в одном организме. Соматическая мутационная изменчивость проявляется заметней тогда, когда она начинается на ранних стадиях. Если мутация случается на первых стадиях дробления зиготы, то возникнет больше клеточных линий с отличными друг от друга генотипами. Соответственно, уже больше клеток будет нести мутацию, такие организмы называются мозаичными.
Уровни наследственных структур
Мутационная изменчивость проявляется в наследственных структурах, отличающихся разным уровнем организации. Мутации могут происходить на генном, хромосомном и геномном уровнях. В зависимости от этого изменяются и виды мутационной изменчивости.
Генные изменения затрагивают структуру ДНК, в результате чего она меняется на молекулярном уровне. Такие изменения в некоторых случаях никак не влияют на жизнеспособность белка, т.е. функции никак не меняются. Но в других случаях могут происходить дефектные образования, что уже прекращает способность белка выполнять свою функцию.
Мутации на хромосомном уровне уже несут более серьёзную угрозу, потому что они влияют на формирование хромосомных болезней. Результатом такой изменчивости являются изменения в структуре хромосом, а здесь уже задействовано сразу несколько генов. Из-за этого может изменяться обычный диплоидный набор, что в свою очередь может в целом повлиять и на ДНК.
Геномные мутации также как и хромосомные могут стать причиной формирования хромосомной болезни. Примеры мутационной изменчивости на этом уровне - анеуплоидия и полиплоидия. Это увеличение или уменьшение числа хромосом, которые для человека чаще всего оказываются летальными.
К геномным мутациям относится трисомия, означающая наличие трёх гомологических хромосомы в кариотипе (увеличение количества). Такой отклонение приводит к формированию синдрома Эдвардса и синдрома Дауна. Моносомия означает наличие только одной из двух гомологических хромосом (уменьшение количества), что практически исключает нормальное развитие эмбриона.
Причиной возникновения подобных явлений становятся нарушения на разных стадиях развития половых клеток. Происходит это в результате анафазного отставания - гомологические хромосомы при делении клетки движутся к полюсам, и одна из них может отставать. Также существует понятие "нерасхождение", когда хромосомы не смогли разделиться на стадии митоза или мейоза. Результатом этого становится проявление нарушений разной степени тяжести. Изучение данного явления поможет разгадать механизмы и, вероятно, даст возможность предсказывать и влиять на эти процессы.
У растений:


Мутации окраски у фиалки

Цветки: а — диплоидного; б — тетраплоидного растения лилии

Соматические мутации, вызванные у растений ионизирующей радиацией (рентгеновские или гамма-лучи): появление белой окраски в красных цветках табака (1) и двух сортов львиного зева (2 и 3); на рис. 3 (слева) — нормальный цветок, справа — мутировавший после облучения.

Мутантные формы ячменя: поздняя полегающая (слева) и ранняя неполегающая (справа).
У животных:


сросшиеся черепашата козленок с n-количеством конечностей


жаба с n-количеством конечностей змея с двумя головами

3. Цитоплазматическое наследование
Участие цитоплазмы в наследовании признаков сопряжено как с действием через нее ядерных генов, так и с работой внеядерных генов, расположенных в ДНК органоидов. Роль материнской цитоплазмы в наследовании признаков, кодируемых ядерными генами. Хорошо известно, что у прудовой улитки бывают как право-, так и левозакрученные раковины. Этот признак формируется действием генов матери на свойства цитоплазмы яйцеклетки. Направление закру- чивания раковины определяется парой аллельных генов Dd. Гомозиготы dd закручены в левую сторону, a DD — в правую. Рассмотрим варианты скрещивания (♀ – женские особи, ♂ – мужские особи):
Р ♀ DD x ♂ dd ♀ dd x ♂DD
F1 Dd Dd.
Генотипы потомства совершенно одинаковы, но в первом случае раковины будут правозакрученными, а втором – левозакрученными, в соответствии с генотипом матери. В чем же дело? Как показали эмбриональные исследования, направление закручивания определяется расположением бластомеров при дроблении яйцеклетки. В какую сторону смещаются по спирали бластомеры каждого следующего ряда, такой и сформируется раковина. Направление смещения бластомеров задается ориентацией митотического веретена деления в процессе дробления, то есть –свойствами материнской цитоплазмы.
Сперматозоиды почти не имеют цитоплазмы, и зигота улитки наследует ее от яйцеклетки. В результате фенотип прудовиков определяется не их собственным генотипом, а генотипом матери. Наследование признаков, кодируемых внеядерными генами. Митохондрии и пластиды содержат небольшие кольцевые молекулы ДНК, которые, как и ядерные, обладают способностью к репликации. Эти органоиды воспроизводятся делением независимо от деления клетки. Их гены кодируют ряд признаков. К числу таких признаков относится пёстролистость у ночной красавицы и львиного зева.
Часть хлоропластов этих растений в результате случайных нарушений ДНК теряет способность синтезировать хлорофилл, поэтому листья становятся пестрыми. Если такие хлоропласты оказываются в Цитоплазме яйцеклетки, то передаются по наследству. Неудивительно, что наследование пестролистости не укладывайся в закономерности, открытые Менделем. Цитоплазматическое наследование признаков у многоклеточных организмов, размножающихся половым путем, осуществляется только по материнской линии по причине почти полного отсутствия цитоплазмы в мужских гаметах.
Свойства цитоплазмы, как и ДНК органоидов, определяют лишь небольшую часть признаков, поэтому цитоплазматическое наследование играет второстепенную роль по сравнению с хромосомной наследственности.


Дифенбахия с бархатными пятнистыми листьями Бегония пятнистая


Хоста яснотка пятнистая


Медуница садовая лист расторопши пятнистой

Хлорофитум хохлатый
4. Закон гомологических рядов
Гомологических рядов закон изменчивости, разработанный советским учёным Н. И. Вавиловым закон, устанавливающий параллелизм в изменчивости организмов. Ещё Ч. Дарвин (1859-68) обратил внимание на далеко идущий параллелизм в изменчивости близких видов и родов животных и растений. В 19 и начале 20 вв. ряд ботаников и зоологов (например, франц. учёный М. Дюваль-Жув, 1865; швейцарский миколог Э. Фишер, 1896; нем. ботаник Э. Цедербауэр, 1907, 1927; рус, зоолог В. М. Шимкевич, 1906, 1921, и др.) специально изучали параллельную изменчивость разных видов растений и животных. Советский генетик Ю. А. Филипченко подытожил (1922) ряд таких, преимущественно зоологических, данных в статье о параллелизме изменчивости в живой природе, который он объяснял систематической и филогенетической близостью родов и видов, входивших в изучаемые группы.
Лишь Н. И. Вавилов подошёл к проблеме параллелизма в изменчивости близких видов и родов с генетических позиций и на основе сравнительного изучения обширнейшего мирового материала (в природных условиях, культурах и в опытах) по изменчивости ряда семейств растений, богатых хорошо изученными культурными видами, - главным образом злаков. Это позволило ему в 1920 на 3-м Всероссийском съезде селекционеров в Саратове выступить с докладом «Закон гомологических рядов в наследственной изменчивости». Н. И. Вавилов показал, что если все известные у наиболее изученного в данной группе вида вариации расположить в определённом порядке в таблицу, то можно обнаружить и у других видов почти все те же вариации изменчивости признаков. Более того, по мере развития исследований видов, входящих в данную группу, «пустые» места в таблице заполняются и параллелизм в изменчивости близких видов становится всё более полным. Принципиально сходный, но слабее выраженный параллелизм характеризует изменчивость различных родов в пределах семейства, и ещё менее полный - различных семейств в пределах группы более высокого ранга.
Таким образом, Гомологических рядов закон сводится к следующему: близкие виды благодаря большому сходству их генотипов (почти идентичные наборы генов) обладают сходной потенциальной наследственной изменчивостью (сходные мутации одинаковых генов); по мере эволюционно-филогенетического удаления изучаемых групп (таксонов), в связи с появляющимися генотипическими различиями параллелизм наследственной изменчивости становится менее полным. Следовательно, в основе параллелизмов в наследственной изменчивости лежат мутации гомологичных генов и участков генотипов у представителей различных таксонов, то есть действительно гомологичная наследственная изменчивость. Однако и в пределах одного и того же вида внешне сходные признаки могут вызываться мутациями разных генов; такие фенотипические параллельные мутации различных генов могут, конечно, возникать и у разных, но достаточно близких видов. Н. И. Вавилов подчёркивал, что Гомологических рядов закон неизбежно обнимает и такую, в генетическом смысле не строго гомологичную, фенотипически же параллельную изменчивость.
После 1920 представители школы Н. И. Вавилова в СССР, а также ботаники и селекционеры зарубежных стран накопили огромный фактический материал, подтверждающий всеобщность Гомологических рядов закон
Сначала исследования касались в основном морфологических признаков; затем они были распространены на биологические, физиологические и биохимические свойства. Многочисленные подтверждения Гомологических рядов закон были получены на простейших, низших растениях, большом числе семейств высших растений и на животных.
Гомологических рядов закон отражает всеобщее и фундаментальное явление в живой природе. Он имеет огромное практическое значение в растениеводстве и селекции, а также в животноводстве. На основе этого закона растениеводы и животноводы могут целенаправленно искать и находить нужные признаки и варианты у различных видов в почти бесконечном мировом многообразии форм как культурных растений и домашних животных, так и у их диких родичей. Эти поиски, особенно среди культурных растений и их диких предков, значительно облегчаются учением Н. И. Вавилова (1926 и др.) о центрах происхождения культурных растений и его работами (1927, 1928, 1930) о географических закономерностях в распределении генов культурных растений. Гомологических рядов закон Н. И. Вавилова уже с 30-х гг. 20 в. явился мощным стимулятором целенаправленной селекции, создания новых сортов культурных растений и разработки научных основ интродукции и акклиматизации. Гомологических рядов закон играет всё большую роль в изучении механизмов эволюционного процесса, в истолковании ряда биогеографических явлений и в разработке основ современной систематики низших таксонов.
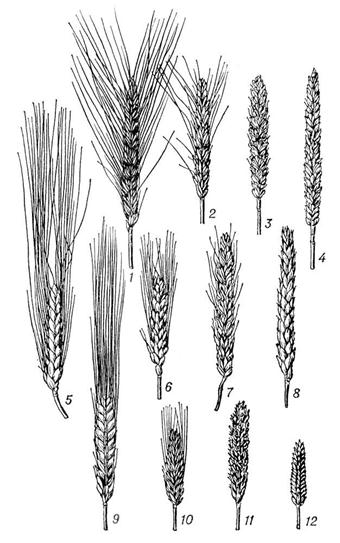
Параллельная изменчивость остистости колоса у мягкой пшеницы (1-4), у твёрдой пшеницы (5-8) и у шестирядного ячменя (9-12) (по Н. И. Вавилову).
5.Центры происхождения культурных растений.
Все культурные растения, которыми сегодня питается человечество, произошли от диких видов растений, которые древний человек использовал в пищу. Потом люди заметили, что эти растения значительно выгоднее выращивать самим. Тогда зародилось земледелие. Момент возникновения земледелия - это коренной перелом в истории человеческого общества. Возделывание земли позволило человеку многократно увеличить количество пищи, ведь теперь он не только охотился и собирал, что мог найти в природе, но начал сам производить еду. Этот момент носит также название Неолитическая революция. Раньше всего она началась на Ближнем Востоке - около 10 тысяч лет назад, позднее всего пришла в Америку. Однако, многие народы Земли, такие как американские индейцы, пигмеи или аборигены Австралии и по сей день не перешли к земледелию, предпочитая оставаться охотниками и собирателями также, как поступали их предки ещё десятки тысяч лет назад.
Современные растения и их плоды, которые мы употребляем в пищу уже совсем не те, какими они были тысячи лет назад, когда их впервые обнаружил человек. Урожай за урожаем он отбирал наиболее совершенные растения, так как заметил, что их свойства передаются растениям, выросшим из отобранных семян. Так сначала неосознанно, а потом и намеренно тысячелетиями проводился искусственный отбор и селекция - выведение новых сортов растений. В последние десятилетия люди научились менять растения на генетическом уровне без отбора, а посредством современных технологий генной инженерии. Растения с измененными генами называются генно-модифицированными. Они имеют значительно лучшие показатели по урожайности и приспособленности к неблагоприятным факторам среды, чем их предшественники, выведенные обычной селекцией. Однако, на сколько безопасны генно-модифицированные растения при употреблении в пищу, ещё только предстоит выяснить.
Разумеется, разные растения произошли в разных регионах планеты. Изучением географии их происхождения учёные начали заниматься после открытия Чарльзом Дарвином законов эволюции и отбора. Дарвин выдвинул идею о том, что все виды имели центры происхождения, где они возникли впервые. В 1883 А. Декандоль опубликовал труд, в котором установил географические области начального происхождения главнейших культурных растений. Однако, наиболее полную и целостную теорию центров происхождения видов культурных растений сформулировал Николай Иванович Вавилов (на фото) в ходе исследований в 20-30-е годы прошлого века. Для этого ему пришлось объехать практически все сельскохозяйственные районы Земли - в Америке, Африке и Евразии. В ходе своих экспедиций Вавилов собрал богатейшую коллекцию культурных растений, нашел родственные связи между ними, предсказал возможные для выведения ранее неизвестные, но заложенные генетически, свойства этих культур. К сожалению, наука генетика, без которой невозможно было бы ни совершить все эти открытия, ни развивать продуктивность сельского хозяйства, в Советском Союзе была признана ложной и идеологически враждебной. Так как она утверждала, что свойства живых существ, а в том числе и человека, определяет наследственность, что противоречило идеям всеобщего равенства, пропагандируемого коммунистическим режимом. Н.И. Вавилов, как многие другие советские ученые тех времен, в 1940-м году был репрессирован и в 1943 году умер в тюрьме.
В ходе исследования Н.И. Вавилова было выявлено 7 основных географических центров происхождения культурных растений.
Южноазиатский тропический центр (около 33 % от общего числа видов культурных растений). (огурец, лимон, джут, манго, рис, банан, кокосовая пальма, черный перец)
Восточноазиатский центр (20 % культурных растений). (просо, соя, редька, грецкий орех, мандарин, хурма, бамбук, женьшень)
Юго-Западноазиатский центр (4 % культурных растений). (пшеница, ячмень, рожь, слива, фундук, финиковая пальма)
Средиземноморский центр (примерно 11 % видов культурных растений). (овёс, лён, лавр, виноград, капуста, кабачок, петрушка, сельдерей, горох, бобы, морковь, свёкла, редис, мята, тмин, хрен, укроп)
Эфиопский центр (около 4 % культурных растений). (кофе, сорго, хлопчатник, кунжут, арбуз)
Центральноамериканский центр (примерно 10 %). (кукуруза, фасоль, тыква, какао, перец, подсолнечник, табак, топинамбур, папайя)
Андийский (Южноамериканский) центр (около 8 %) (картофель, помидор, ананас, гевея, арахис)
Южноамериканский центр
По Н. И. Вавилову: Новый Свет (Перу), 1926; Южноамериканский (Перуано-Эквадоро-Боливийский) центр с тремя очагами - собственно Андийским, Чилоанским и Бразильско-Парагвайским, 1935; Андийский центр (Южноамериканский) с тремя очагами - собственно Андийским, Чилоанским и Боготанским (Боготским), 1939, 1962. По Е. Н. Синской: Новосветская область (Южноамериканская подобласть), 1969. По П. М. Жуковскому: «Южноамериканский генцентр, 1971. По А. И. Купцову: Перуанский очаг, 1975. По Тахтаджяну: Андийская область, 1978.
Южноамериканский генцентр включает прибрежные и субтропические области Перу, Эквадора, Боливии, Бразилии, Чили, Парагвая и других стран этого континента, а также прилегающих к ним островов.
В этом генцентре - одно эндемичное семейство и несколько сот эндемичных родов (20). Н. И. Вавилов, характеризуя центр, назвал 59 основных эндемичных культивируемых видов и их диких сородичей, причем только картофеля среди них - 16 видов, а также зерновые, плодовые, овощные и другие растения. П. М. Жуковский привел для генцентра 69 основных видов. Этот генцентр - родина культурного картофеля и других клубненосных растений. Здесь возникли первичные центры томата, арахиса, длинноволокнистого хлопчатника, ананаса, хинного дерева и пр. (13).
Средиземноморский центр
По Н. И. Вавилову: центр Средиземноморского побережья, 1926; Средиземноморский очаг, 1935; Средиземноморский центр, 1939, 1962. По Е. Н. Синской: Древнесредиземноморская область (с подобластью собственно Средиземноморье), 1969. По П. М. Жуковскому: Средиземноморский генцентр, 1971. По А. И. Купцову: Средиземноморский очаг, 1975. По А. Л. Тахтаджяну: Средиземноморская область, 1978.
Средиземноморский генцентр - один из древнейших центров растениеводства (13). Его растительный мир формировался на стыке бореальных и тропических флор; он включает одно эндемичное семейство, число же эндемичных родов в этом генцентре, вероятно, достигает 150 (20). Человек использует виды следующих эндемичных родов: Argania (масличные и кормовые), Ceratonia (пищевые и кормовые), Facto-rovskya (близок к Trigonella), Lygaeum (зерновые), Bonannia (плодовые), Majorana (эфирно-масличные), Fedia (пищевые), Prasium (дубильные, лекарственные и пряные), Rosmarinus (пряные, эфирно-масличные, лекарственные), Chamaerops (волокнистые) и др.
Н. И. Вавилов (7) привел в перечне культурных растений этого генцентра 84 вида. В частности, он считал Средиземноморский генцентр родиной маслины и рожкового дерева. П. М. Жуковский (13) характеризует его 65 видами культурных растений и их дикорастущих сородичей.
К автохтонным культурным растениям принадлежат крупносеменые овсы (Avena byzantina), лен, мак, белая горчица, маслины, капуста, морковь, свекла, лук, чеснок [15]. Многие культурные растения, например лен, ячмень, бобы, нут, отличаются крупнозерностью и крупноплодностью (7).
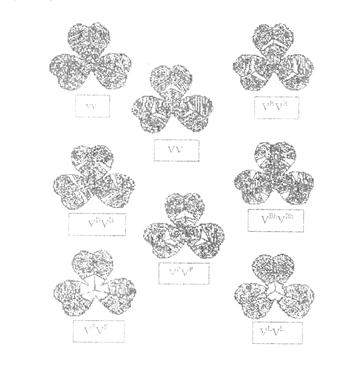
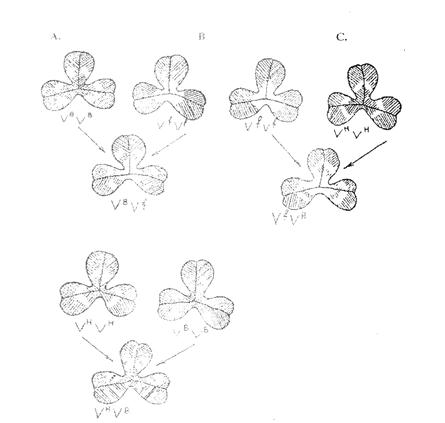
6. Изучение изменчивости седых пятен клевера
Фенотипы, обусловленные мутациями V-локуса у клевера ползучего
| Аллель | Фенотип |
| v | Пятно отсутствует |
| V | Полное пятно |
| VH | Полное пятно, высокое |
| VB | Разорванное пятно |
| VBh | Разорванное высокое |
| VP | Центральная верхняя точка |
| VF | Большое сплошное пятно у основания |
| VL | Низкое треугольное пятно у основания |

Дата публикования: 2014-11-04; Прочитано: 1218 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
