 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Глава 4. Механизмы генерации потенциала действия
|
|
Экспериментальной базой для создания модели генерации потенциала действия явились результаты опытов по разделению ионных токов возбужденного аксона
Для разделения токов использовали блокатор натриевого тока - тетродотоксин (ТТХ) и блокатор калиевого тока - тетраэтиламмоний (ТЭА).
Измерение входящих и выходящих токов проводилось в режиме фиксации потенциала. При введении в раствор тетродотоксина регистрировали временную зависимость выходящего тока К+, IK(t) При действии на мембрану тетрааэтиламмонием регистрировали временную зависимость входящего тока Na+, INa (t) при тех же значениях деполяризующего потенциала, что и для калиевого тока.
В серии опытов на аксоне кальмара было показано:
1) образование потенциала действия связано с переносом ионов Na+ и К+ через мембрану;
2) проводимость мембраны для этих ионов меняется в зависимости от величины мембранного потенциала и времени:
gNa(φM,t), gK(φM,t).
В дальнейшем Ходжкин и Хаксли предложили математическую модель, которая описывала изменения проводимостей, а следовательно, и токов ионов Na+ и К+ через мембрану в процессе возбуждения.
Основными постулатами этой модели являются:
1) в мембране существуют отдельные каналы для переноса ионов Na+и К+;
2) во внутренней структуре мембраны существуют некоторые заряженные частицы, управляющие проводимостью каналов. В зависимости от величины напряженности приложенного электрического поля эти гипотетические частицы могут передвигаться в мембране, и тем самым увеличивать или уменьшать потоки ионов Na+ и К+ через каналы.
Предполагается, что ионы калия могут проходить через канал, если к его участку под действием электрического поля подойдут одновременно четыре однозарядные частицы. Обозначим n - вероятность подхода одной такой частицы. Тогда проводимость ионов калия:
gK=gK∙n4
где gK - максимальная проводимость канала для ионов К+. Четвертая степень при n определялась эмпирически. Величина n4 объяснялась как вероятность нахождения одновременно четырех активирующих частиц в некотором определенном участке мембраны.
Изменение проводимости для ионов Na+ описывалось более сложным выражением. Для натриевого канала предполагалось, что он открывается, если одновременно в данный участок попадают три активирующие частицы и удаляется одна блокирующая. Тогда, обозначив m - вероятность прихода активирующей частицы, a h- вероятность удаления блокирующей, получаем:
gNa=gNa∙m3h
где gNa - максимальная проводимость канала для ионов Na+.
Здесь введены два типа частиц, активирующие и блокирующие, так как натриевый ток в условиях фиксированного потенциала сначала нарастает до максимума - активация, а затем уменьшается до 0 - инактивация. Степени при m и h также подбирались эмпирически, чтобы наилучшим образом описать кинетику токов. Численные значения n, m и h имеют смысл вероятности нахождения соответствующей частицы в данном месте канала, а величины их могут меняться от 0 (отсутствие частицы) до 1 (нахождение ее в заданном месте.
Физическая интерпретация модели Ходжкина-Хаксли требовала наличия внутри мембраны некоторых заряженных частиц, причем эти частицы должны передвигаться в зависимости от внешнего электрического поля. Таким образом, для подтверждения второго постулата модели необходимо было зарегистрировать перемещения заряженных частиц внутри мембраны при изменении мембранного потенциала, то есть зарегистрироватъ так называемые воротные токи. Трудное обнаружения воротных токов заключалась в том, что активирующих частиц внутри мембраны очень мало и, следовав но, мало значение воротного тока по сравнению с ионными токами, проходящими через мембрану.
Для обнаружения воротных токов с помощью блокаторов ТТХ и ТЭА, а также заменой ионов Na+ в наружном растворе на ионы триса, исключали ионные токи; затем ступеньками меняли напряжение на мембране и регистрировали появление воротного тока натриевого канала, который оказался в 103 слабее натриевого тока.
Изменение во времени воротного тока в аксоне кальмара было взаимосвязано с изменением натриевого тока. Таким образом: на опыте было показано существование воротных токов, предсказанных в модели Ходжкина-Хаксли.
Ионные каналы клеточных мембран
Модель возбудимой мембраны по теории Ходжкина-Хаксли предполагает регулируемый перенос ионов через мембрану. Однако непосредственный переход иона через липидный бислой весьма затруднен. Поэтому величина коэффициента распределения К в формулах очень мала, а следовательно, был бы мал и поток ионов, если бы ион переходил непосредственно через липидную фазу мембраны. Таким образом, непосредственный перенос ионов через липидный бислой только за счет диффузии маловероятен.
Можно предположить, что в мембране должны быть некоторые специальные структуры - проводящие ионы. Такие структуры были найдены и названы ионными каналами. Подобные каналы выделены из различных объектов: плазматической мембраны клеток, постсинаптической мембраны мышечных клеток и других объектов. Известны также ионные каналы, образованные антибиотиками.
Основные свойства ионных каналов:
1) селективность;
2) независимость работы отдельных каналов;
3) дискретный характер проводимости;
4) зависимость параметров каналов от мембранного потенциала.
Рассмотрим их по порядку.
1. Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа.
Еще в первых опытах на аксоне кальмара было обнаружено что ионы Na+ и К+ по-разному влияют на мембранный потенциал. Ионы К+ меняют потенциал покоя, а ионы Na+ - потенциал действия. В модели Ходжкина-Хаксли это описывается путем введения независимых калиевых и натриевых ионных каналов. Предполагалось, что первые пропускают только ионы К+, а вторые - только ионы Na+.
Измерения показали, что ионные каналы обладают абсолютной селективностью по отношению к катионам (катион-селективные каналы) либо к анионам (анион-селективные каналы). В то же время через катион-селективные каналы способны проходить различные катионы различных химических элементов, но проводимость мембраны для неосновного иона, а значит, и ток через нее, будет существенно ниже, например, для Na+-кaнала калиевый ток через него будет в 20 раз меньше. Способность ионного канала пропускать различные ионы называется относительной селективностью и характеризуется рядом селективности - соотношением проводимостей канала для разных ионов, взятых при одной концентрации. При этом для основного иона селективность принимают за 1. Например, для Na+-канала этот ряд имеет вид:
Na+:K+= 1:0,05.
2. Независимость работы отдельных каналов. Прохождение тока через отдельный ионный канал не зависит от того, идет ли ток через другие каналы. Например, К+-каналы могут быть включены или выключены, но ток через Nа+-каналы не меняется. Влияние каналов друг на друга происходит опосредованно: изменение проницаемостей каких-либо каналов (например, натриевых) меняет мембранный потенциал, а уже он влияет на проводимости прочих ионных каналов.
3. Дискретный характер проводимости ионных каналов. Ионные каналы представляют собой субъединичный комплекс белков, пронизывающий мембрану. В центре его существует трубка, сквозь которую могут проходить ионы. Количество ионных каналов на 1 мкм2 поверхности мембраны определяли с помощью радиоактивномеченного блокатора натриевых каналов - тетродотоксина. Известно, что одна молекула ТТХ связывается только с одним каналом. Тогда измерение радиоактивности образца с известной площадью позволило показать, что на 1 мкм2 аксона кальмара находится около 500 натриевых каналов. Те трансмембранные токи, которые измеряют в обычных экспериментах, например, на аксоне кальмара длиной 1 см и диаметром 1 мм, обусловлены суммарным ответом (изменением проводимости) 500 • 3 • 107 - 1010 ионных каналов. Для такого ответа характерно плавное во времени изменение проводимости. Ответ одиночного ионного канала меняется во времени принципиально иным образом: дискретно и для Na+-каналов, и для К+-, и для Са2+-каналов.
Результаты экспериментов, выполненных на различных ионных каналах, показали, что проводимость ионного канала дискретна и он может находиться в двух состояниях: открытом или закрытом. Переходы между состояниями происходят в случайные моменты времени и подчиняются статистическим закономерностям. Нельзя сказать, что данный ионный канал откроется именно в этот момент времени. Можно лишь сделать утверждение о вероятности открывания канала в определенном интервале времени.
Рассмотрим токи через одиночные Ка+-каналы.
Канал за время одного такого деполяризующего сдвига открывался лишь один раз на время tи, которое будем называть временем открытого состояния канала.
Среднее значение tи для Na+-канала ≈ 0,7 мс (от 0,3 до 1,5 мс).
Одиночный канал может открыться раньше (1-й опыт) или позже (N-й опыт). Время, в течение которого вероятность открывания отдельного канала велика, будем называть временем жизни каналов: TNa, ТСа. Для натриевых каналов TNa≈ 2 мс.
Таким образом, процесс открытия натриевых каналов - процесс стохастический: сдвиг φм выше порогового значения увеличивает вероятность открывания каналов, то есть идет процесс их активации. По прошествии времени жизни каналов TNa вероятность их открывания падает до нуля и этот процесс называется инактивацией Na+-токa.
Несмотря на то, что ток через каждый ионный канал меняется скачком, зависимость суммарного трансмембранного тока во времени плавная Этот феномен можно объяснить, используя методы статистической физики.
Суммарный ток I через N одиночных ионных каналов:
I=  ,
,
где in – ток через n-й канал.
Среднее значение I суммарного тока в случае одинаковых каналов определяется средним током i в каждом канале:
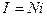
Относительная флуктуация тока в одиночном канале велика:
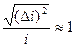
При больших N относительные флуктуации ничтожны. Для совокупности N = 1010 ионных каналов, расположенных на участке аксона кальмара, флуктуация тока составляет 10-5 (0,001 %) от среднего значения тока через мембрану, то есть флуктуации тока при измерениях в этом случае практически не заметны. Для маленьких клеток, в которых может быть порядка 103 ионных каналов, относительные флуктуации более существенны.
Токи одиночных К+-каналов имеют амплитуду до 2 пА, а среднее время открытого состояния ta ≈ 5 мс. Однако за это время канал может несколько раз открыться и закрыться на короткое время, то есть могут происходить осцилляции тока. В отличие от натриевых, К+-каналы не инактивируются, пока φм выше порогового значения. Отдельные каналы во время деполяризации могут открываться по нескольку раз.
Токи одиночных Са2+-каналов кардиомиоцитов имеют более сложный характер по сравнению с Na+- и К+-токами аксонов. Во время последовательных скачков деполяризации в 70 % случаев Са2+-канал открывается на время ~ 1 мс; затем через каждые 0,2 мс он закрывается и вновь открывается и пропускает ток с амплитудой импульса ≈ 1 пА. Такой процесс активации Са2+-тока длится около 130 - 200 мс, а затем наступает инактивация Са2+-тока. В 30 % скачков деполяризаций кальциевый канал остается закрытым.
4. Зависимость параметров канала от мембранного потенциала Ионные каналы нервных волокон чувствительны к мембранному потенциалу, например натриевый и калиевый каналы аксона кальмара. Это проявляется в том, что после начала деполяризации мембраны соответствующие токи начинают изменяться. Ион-селективный канал имеет сенсор - некоторый элемент своей конструкции, чувствительный к действию электрического поля. При изменении мембранного потенциала меняется величина действующей на него силы, в результате эта часть ионного канала перемещается и меняет вероятность открывания или закрывания ворот - своеобразных заслонок, действующих по закону "все или ничего". Экспериментально показано, что под действием деполяризации мембраны увеличивается вероятность перехода натриевого канала в проводящее состояние. Скачок напряжения на мембране, приводит к тому, что большое число каналов открывается. Через них проходит больше зарядов, а значит, в среднем, протекает больший ток. Существенно, что процесс роста проводимости канала определяется увеличением вероятности перехода канала в открытое состояние, а не увеличением диаметра открытого канала. Таково современное представление о механизме прохождения тока через одиночный канал.
Плавные кинетические кривые токов, регистрируемых при электрических измерениях на больших мембранах, получаются вследствие суммации многих скачкообразных токов, протекающих через отдельные каналы. Их суммирование, как показано выше, резко уменьшает флуктуации и дает достаточно гладкие зависимости трансмембранного тока от времени.
Ионные каналы могут быть чувствительны и к другим физическим воздействиям: механическим деформациям, связыванию химических веществ и т.д. В этом случае они являются структурной основой, соответственно, механорецепторов, хеморецепторов и т.д.
Изучение ионных каналов в мембранах есть одна из важнейших задач современной биофизики.
Дата публикования: 2014-11-04; Прочитано: 1912 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
