 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Материалы и методы
|
|
Введение
Теломеры представляют собой повторяющиеся последовательности ДНК (у млекопитающих это двуцепочный участок, состоящий из повторов TTAGGG, и 3’-выступающий 150-200 нуклеотидный участок G-цепи) и связанные с ними специальные белки, которые защищают концы хромосом от деградации и системы репарации двуцепочечных разрывов. Длина теломер определяет, какое количество делений клетка может совершить до своей гибели, то есть пролиферативный потенциал клетки. Теломераза – фермент, поддерживающий длину теломер. При повышенной активности теломеразы длина теломер поддерживается на постоянном уровне, благодаря чему клетка может совершать бесконечное число делений. Наибольшая активность теломеразы наблюдается в раковых клетках, следовательно, надо каким-нибудь образом ингибировать теломеразную активность. Было установлено, что теломерная последовательность, богатая гуанинами, может образовывать структуры, способные ингибировать теломеразу. Это, так называемые, G-квадруплексы, состоящие из гуаниновых тетрад, уложенных в стопку друг на друга. К сожалению, существующие методы анализа стабилизации квадруплексов имеют ряд недостатков: достаточно сложны, либо требуют работы с радиоактивной меткой и др. Известно, что агрегация наночастиц золота приводит к быстрому изменению цвета раствора, что может быть хорошим и быстрым методом детекции. Принцип работы основывается на насаживании олигонуклиотидов, которые способны образовывать квадруплексы (т.е.богатые гуанином последовательности) на наночастицы, тем самым защищая частицы от агрегации.[9]
Лит.Обзор
G-квадруплекс (или G4 DNA) – это стабильная четырех цепочная структура, образующаяся на гуанин богатых участках ДНК последовательности; было определенно, что формирование таких структур в промоторах некоторых генов (например: MYC, KRAS, PDGFA и INS) существенно влияет на уровень транскрипции генов. Все большее число G4 DNA формирований идентифицировано и охарактеризовано в функциональных областях (промоторах и усилителях) рост-связанных генов, включая KIT, HIF1A, VEGFA, BCL2, RB1. G-квадруплексы распространенны по всему геному человека, что повышает возможность того, что они могут играть общую роль в регуляторном сигнале. Так же возможность формирования квадруплекса в участке транскрипции генов коррелирует с функциональными предпочтениями, предполагают, что гены с одинаковыми или схожими функциями могут корегулироваться в зависимости от присутствия G4 DNA сигнала. [1]
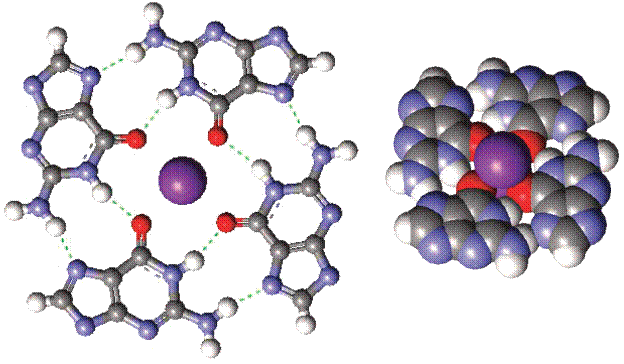
Рис 1: G-квартет, образованный четырьмя гуанинами, связанными между собой водородными связями, включая центральный катион калия, молекулы сахара убраны для краткости.
G-квадруплекс представляет собой вторичную структуру нуклеиновой кислоты, образованную четырьмя гуанинами (рис 1); формируется в присутствии катионов (калия). Каждый квартет состоит из четырёх гуанинов, связанных вместе в упорядоченный цикл восьмью водородными связями… В обычном ДНК-дуплексе, четыре нуклеотидных основания образуют две конкретные пары: аденин с тимином, и гуанин с цитозином. Однако эти пары отличаются стабильностью, так пара гуанин-цитозин более стабильна. Это можно объяснить образованием трех водородных связей, в среднем по полторы связи на основание, тогда как у аденина с тимином – две пары, то есть одна связь на основание. В квадруплексе образуются восемь водородных связей между четырьмя основаниями, то есть по две связи на основание в среднем. Следовательно, структура квадруплекса должна быть более стабильной, нежели G-C-богатые дуплексы ДНК. Хотя этот расчет является очень упрощенным, вывод верен: квадруплексы являются очень стабильной структурой, при этом многие стабильны при более 60°С в физиологических концентрациях соли. Поэтому можно предположить, что они могут образовываться внутри клеток и быть при этом очень стабильными. [2]
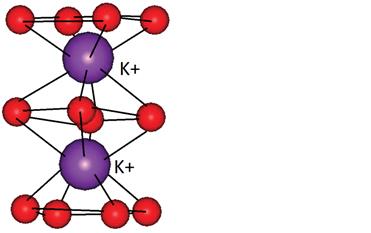
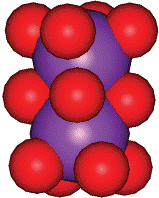
Рис 2: Катион калия связывает восемь карбонильных атом кислорода с длиной связи 2.73А.
Связывание в квартете происходит через два атома азота (N1и N2) с одним основанием, а с другим – через седьмой атом азота (N7) и шестой атом кислорода (О6)… две и более таких структур наложенных друг на друга и связанных между собой через фосфатные группы, образуют G- квадруплекс (рис 2). Атомы кислорода (О6) формируют центральный канал. Присутствие катиона в центральном канале между парой тетрады позволяет стабилизировать структуру. В зависимости от расположения катиона и атомов О6 и происходит стабилизация. Ион, который эффективно координирует, увеличивает стабильность. Это наиболее хорошо видно в экспериментах со связыванием при участии катионов натрия и калия. Увеличение концентрации моновалентного катиона немного увеличивают диссоциацию внутримолекулярных квадруплексов, но значительно увеличивают скорость их перекрутки. Было установлено, что десятикратное увеличение концентрации натрия увеличивает скорость свертки для последовательности d(ТG4Т) в 200 раз.[2] Этот эффект увеличивается при добавлении катионов калия, которые имеют идеальные характеристики для стабилизации О6. Они располагается на расстоянии 2.73 А от каждого О6. Ионы натрия располагаются немного ниже или в стороне от центрального положения, ближе к тетраде. Тенденция стабилизации выглядит следующим образом: K>Na>Rb>NH4>Cs>Li. Прибавление бивалентных ионов, например, магния, может способствовать ассоциации квадруплексов, но в то же время и дестабилизировать сформированный квадруплекс, тем самым уменьшая общую стабильность.





Рис 3: Топология закрутки квадруплексов; стрелки указывают направление 5`-3`: а) межмолекулярный и параллельный; б-в) бимолекулярные и антипараллельные; г-д) внутримолекулярные.
Различают тетрамолекулярные, бимолекулярные и внутримолекулярные квадруплексы (Рис 3). Тетрамолекулярные образованы четырьмя отдельными цепочками последовательности, состоящими из непрерывных рядов гуанина. Бимолекулярные формируются двумя цепочками последовательности, в которой ряд гуанина прерывается другими основаниями. Внутримолекулярные квадруплексы формируются одной цепочкой последовательности, в которой четыре ряда гуанина разделены между собой последовательностями других нуклеотидов. В зависимости от того как направлены цепи последовательности в бимолекулярном или внутримолекулярном квадруплексе, структура может принимать одно из ряда топологий с разными конформациями цикла. Основания в нормальной двуцепочечной ДНК находятся исключительно в anti конформации, в то время как гуанины в квадруплексе могут быть как в anti так и в syn конформации (рис 4).
А)  б)
б) 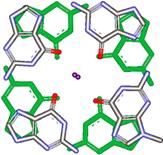 в)
в) 
Рис 4: Связывание гуаниновых тетрад с вариациями расположения цепей в syn и anti положениях: а) anti б) syn-anti в) anti-syn.
Однако если гуанины находятся в одной тетераде и на параллельной пряде, то они должны иметь одинаковые торсиальные углы гликозидной связи. Наоборот, если они находятся на антипараллельной нити, они должны иметь противоположные кручения гликозидных углов. [3] Важную роль в формировании квадруплекса играет нуклеолин. Нуклеолин – это ядрышковый фосфопротеин, который высоко выражается в пролиферации клеток; известный, главным образом, за его роль в биогенезе рибосомы, однако нуклеолин также принимает участие транскрипции, апоптозе и закрутке квадруплексов. Было выявлено, что нуклеолин формирует квадруплекс из одноцепочечной последовательности ДНК и, затем стабилизирует полученную структуру. [5] Для обнаружения квадруплексов используют несколько методов, например: ультрафиолетовое плавление, спектроскопия кругового дихроизма, электрофорез в полиакриламидном геле, FRET, анализ скорости седиментации и метод, основанный на сшивании ДНК и топологии.
Однако могут образовываться структуры, состоящие не только из гуанина. Например, образование i-мотива на цитозин-богатых цепочках с протонированием интерколированных пар C-C. В случае ДНК дуплекса диссоциация цитозин и гуанин богатых цепей требует перекручивания в i-мотив или квадруплекс. В нормальных физиологических условиях клетки гуанин и комплементарные последовательности формируют дуплекс ДНК. Но при низких значениях рН менее 4.5 или при более высоких температурах происходит разъединение и повторное сворачивание цитозин-гуанин богатых цепей с образованием i-мотива для C-богатой и квадруплекса для G-богатой. i-мотив формируется на C-богатой цепи человеческой теломерной последовательности. В этой структуре цепи направлены антипараллельно образуя внутримолекулярный четырех-цепевой квадруплекс. Цитозиновые основания связанные водородными связями в параллельные цепи формируют два широких канала и два узких канала.[4] Последовательности содержащие не только гуанин и цитозин тоже могут образовывать тетрады соединяясь вместе. Человеческая структура квадруплекса содержит один такой пример. Сформированный из двух теломерных повторов (ТАGGGТТАGGGТ)2 повторяет тимин и аденин на 5`-конце. Водородные связи по Уотсону-Крику образуются между аденином и тимином, таким образом получаем антипараллельные цепи. Водородные связи образуются между О6 от тимина и N6 от аденина и стабилизируются катионом натрия. Другой пример можно привести с последовательностью GАGCАGGТ. В данном случае формируется димерный квадруплекс содержащий GCGC, GGGG и неустойчивый АТАТ тетрады. [2]
Теломераза человека – это рибонуклеопротеин, обратный транскриптазе, который катализирует присоединение теломерных повторов (ТТАGGG)n, где Т – тимин, А – аденин, G – гуанин и n – количество повторов на конце хромосомы человека. Этот фермент поддерживает длину теломер. Теломеры представляет собой повторяющиеся последовательности ДНК и связанные с ними специальные белки, которые защищают концы хромосом от деградации и системы репарации двуцепочечных разрывов. У млекопитающих теломеры представляют собой двуцепочечный участок, состоящий из повторов TTAGGG, и 3’-выступающий 150-200 нуклеотидный участок G-цепи. Длина теломер определяет количество возможных делений клетки. Когда теломераза активна, длина теломер поддерживается на постоянном уровне. Клетки в таком случае делятся неограниченно. Такое наблюдается в эмбриональных клеточных линиях, в стволовых клетках активность теломеразы несколько меньше, в соматических клетках активность теломеразы обычно отсутствует. Укорочение теломер приводит к достижению предела Хайфлика (предел Хайфлика – число делений, которое может совершить клетка пока теломераза не достигнет критической длины. В среднем, человеческая клетка может совершить, примерно, 52 деления, после чего она переходит в состояние сенессенса (старение)). Затем, у клеток начинается кризис, заканчивающийся массовой смертью. Уцелевшие клетки перерождаются в раковые, способные к неограниченному делению (поддержание постоянной длины теломеры, за счет активности теломеразы). Человеческая теломерная G-цепь может складываться во внутримолекулярные G-квадруплексные структуры, которые различаются позицией смежной петли; такие структуры будут ингибировать теломеразную активность.
Материалы и методы
В подготовленный раствор с AuNPs (V=25 mL, c=0.04µМ) было добавлено 15 г BSPP, перемешено и оставлено на ночь при комнатной температуре. На следующий день в раствор добавили раствор NaCl (приливается постепенно, постоянно помешивая, пока окраска раствора с красной не станет серо-синей). Далее полученный раствор переносится на центрифугу на 20 минут при 3500об. Таким образом, были осаждены н/ч; верхний слой воды сливается и оставшиеся н/ч растворяются в фосфатном буфере (1mg:1mL H20) до финальной концентрации н/ч 20 µМ. Затем добавляются олигонуклеотиды (mazh-3 5'> TTT TTT TTT TTT AGG GTT AGG G <3' концентрация 208µМ, mazh-4 5'> TTT TTT TTT TTT AGG GTT AGG GTT AGG GTT AGG G <3 концентрация 180,36µМ и pA30 концентрация 156µМ) и полученные растворы оставляют инкубироваться и размешиваться на ночь. В первый раз использовались только нуклеотиды с двумя теломерными повторами Mazh-3. Производилась раститровка на оптимальную концентрацию катионов калия (0.1mМ, 0.3mМ, 0.7mМ и 1М) к количеству олигонуклеотидов на поверхности наночастицы (1/2,1/4,1/16).
После приготовлении растворов н/ч + olig, где концентрация наночастиц была 5мМ, их прокипятили при температуре 95С в течение 3 минут и затем подв.центрифугированию при 13000g в течение 5 минут. Полученные таким образом наночастицы с «развернутыми» олигонуклеотидами использовались в дальнейших опытах.
Дата публикования: 2015-11-01; Прочитано: 654 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
