 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Строение, функции и эволюция пиРНК
|
|
Пока молекулярным биологам были известны только два вида малых РНК — киРНК и микроРНК — основное «предназначение» РНК-интерференции казалось совершенно понятным. Действительно: набор гомологичных коротких РНК и белков у разных организмов осуществляет аналогичные действия; по мере усложнения организмов усложняется и функциональность.
Однако в процессе эволюции природа создала ещё одну, эволюционно самую позднюю и узкоспециализированную систему на основе всё того же удачного принципа РНК-интерференции. Речь идет о пиРНК (piRNA, от Piwi-interaction RNA).
Чем сложнее организован геном, тем более развит и приспособлен организм. Однако увеличение сложности генома имеет и оборотную сторону: сложная генетическая система становится нестабильной. Это ведет к необходимости механизмов, отвечающих за поддержание целостности генома — иначе самопроизвольное «перемешивание» ДНК просто выведет её из строя. Мобильные генетические элементы ( МГЭ ) — один из основных факторов нестабильности генома — представляют собой короткие нестабильные участки, которые могут автономно транскрибироваться и мигрировать по геному. Активация таких мобильных элементов приводит к множественным разрывам ДНК в хромосомах, чреватых летальными последствиями.
Количество МГЭ нелинейно увеличивается с размером генома, и их активность необходимо сдерживать. Для этого животные, уже начиная с кишечнополостных, используют всё тот же феномен РНК-интерференции. Эту функцию также выполняют короткие РНК, однако не те, о которых речь уже шла, а третий их тип — пиРНК.
пиРНК — короткие молекулы длиной в 24–30 нуклеотидов, закодированные в центромерных и теломерных областях хромосомы. Последовательности многих из них комплементарны известным мобильным генетическим элементам, однако есть множество других пиРНК, совпадающих с участками рабочих генов или с фрагментами генома, функции которых неизвестны.
пиРНК (также как и микроРНК) закодированы в обеих цепях геномной ДНК; они весьма разнообразны (до 500 тыс. видов в одном организме). В отличие от киРНК и микроРНК, они образуются одной цепью с характерной особенностью — урацилом (U) на 5’-конце и метилированным 3’-концом. Есть и другие отличия:
1. в отличие от киРНК и микроРНК, они не требуют процессинга Dicer’ом;
2. гены пиРНК активны только в зародышевых клетках (во время эмбриогенеза) и окружающих их эндотелиальных клетках;
3. белковый состав системы пиРНК иной — это эндонуклеазы класса Piwi (Piwi и Aub) и отдельная разновидность Argonaute — Ago3.
Процессинг и активность пиРНК пока достаточно плохо изучены, но уже ясно, что механизм действия совершенно отличается от других коротких РНК — сегодня предложена пинг-понг модель их работы (рис.7).
Главная функция пиРНК — подавление активности МГЭ на уровне транскрипции и трансляции. Считается, что пиРНК активны только во время эмбриогенеза, когда непредсказуемые перетасовки генома особенно опасны и могут привести к гибели зародыша. Это логично — когда иммунная система ещё не заработала, клетки эмбриона нуждаются в какой-нибудь простой, но действенной защите. От внешних патогенов эмбрион надежно защищен плацентой (или оболочкой яйца). Но кроме этого необходима оборона и от эндогенных (внутренних) вирусов, — в первую очередь МГЭ.
Эта роль пиРНК подтверждена опытом — «нокаут» или мутации генов Ago3, Piwi или Aub приводят к серьёзным нарушениям развития (и резкому увеличению числа мутаций в геноме такого организма), а также вызывают бесплодие за счёт нарушения развития половых клеток.
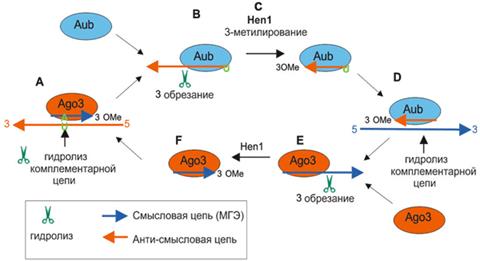
Рисунок 7 А -: Цитоплазматическая часть процессинга пиРНК. Биогенез и активность пиРНК опосредуется семейством эндонуклеаз Piwi (Ago3, Aub, Piwi). Активность пиРНК обеспечивается обеими одноцепочечными молекулами пиРНК — смысловой и анти-смысловой, — каждая из которых ассоциирует со специфической эндонуклеазой Piwi. пиРНК узнает комплементарный участок мРНК транспозона (синяя цепь) и вырезает его. Это не только инактивирует транспозон, но и создает новую пиРНК (связанную с Ago3 с помощью метилирования метилазой Hen1 3’-конца). Такая пиРНК, в свою очередь, узнаёт мРНК с транскриптами кластера предшественников пиРНК (красная цепь) — таким способом цикл замыкается и снова вырабатывается нужная пиРНК [1]
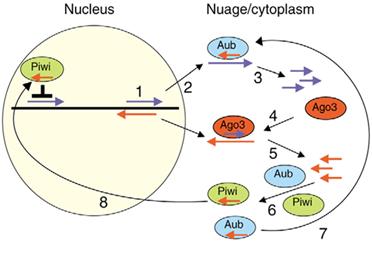
Рисунок 7 Б - пиРНК в ядре. Кроме эндонуклеазы Aub, антисмысловую пиРНК может связывать и эндонуклеаза Piwi. После связывания комплекс мигрирует в ядро, где вызывает деградацию комплементарных транскриптов и перестройку хроматина, вызывающую подавление активности транспозонов [1]
Распространение и эволюция пиРНК. Первые пиРНК обнаруживаются уже у актиний и губок. Растения, видимо, пошли другим путём — белки Piwi у них не обнаружены, а роль «намордника» для транспозонов выполняют эндонуклеаза Ago4 и киРНК.
У высших животных — в том числе и человека — система пиРНК развита очень хорошо, но встретить её можно только в эмбриональных клетках и в околоплодном эндотелии. Почему распространение пиРНК в организме столь ограничено — ещё предстоит узнать. Можно предположить, что, как и любое мощное оружие, пиРНК приносит пользу только в очень специфических условиях (во время развития плода), а во взрослом организме их активность нанесёт больше вреда, чем пользы. Все-таки, число пиРНК на порядок превосходит количество известных белков — и неспецифические эффекты пиРНК в зрелых клетках сложно предсказать.
На рисунке 8 представлена таблица, иллюстрирующая эволюцию белкового аппарата, участвующего в РНК-интерференции. Видно, что у простейших наиболее развита система киРНК (белковые семейства Ago, Dicer), а с усложнением организмов акцент переносится на более специализированные системы — увеличивается число изоформ белков для микроРНК (Drosha, Pasha) и пиРНК (Piwi, Hen1). При этом разнообразие ферментов, опосредующих действие киРНК, уменьшается.
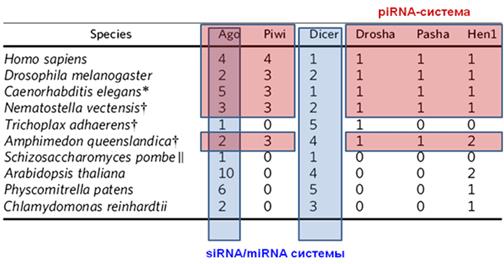
Рисунок 8. Многообразие белков, участвующих в РНК- интерференции (цифры обозначают количество белков каждой группы). Синим цветом подсвечены элементы, характерные для киРНК и микроРНК, а красным — белк и, связанные с пиРНК [1]
Явление РНК-интерференции начали использовать уже простейшие организмы. На основе этого механизма природа создала прототип иммунной системы, а по мере усложнения организмов РНК-интерференция становится незаменимым регулятором активности генома. Два разных механизма плюс три вида малых РНК (см. таблицу) — в результате мы видим тысячи тонких регуляторов различных метаболических и генетических путей. Эта поразительная картина иллюстрирует универсальность и эволюционную адаптацию молекулярных биологических систем. Правда, такая фантастическая сложность говорит скорее о том, что эволюция «слепа» и действует без наперёд утверждённого «генерального плана».
Дата публикования: 2015-10-09; Прочитано: 409 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
