 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Поток генетической информации в клетке. Регуляция экспрессии генов
|
|
Генетическая информация – это «кодовое письмо», содержащее программу о развитии организма. Оно передается от родителей к потомству через половые клетки. О биологической роли этого «кодового письма» Эрвин Шрёдинген еще в 1945 году писал следущее: «Называя структуру хромосомных нитей «кодовым письмом» мы имеем в ввиду, что всепроникающий разум может по этой структуре предсказать, разовьется ли данное яйцо в подходящих условиях в черного петуха или пёструю курицу, в муху или растение кукурузы, жука, мышь или в женщину…» Но это термин «кодовое письмо», конечно, слишьком узок. Хромосомные структуры служат, кроме того, и инструментом, осуществляющим то развитие, которое они же предпологают. Они и кодекс законов и исполнительная сила, или (используя другую терминологию) они и архитектурный проект, и строительная бригада в одно и тоже время.
9.1. Центральная догма (основной постулат) молекулярной биологии
В результате фундаментальных исследований молекулярных основ наследственности сформировалось представление об ауто- и гетеро- синтетической функции гена. Согласно этому представлению передача генетической информации происходит от ДНК к ДНК при репликации ДНК (аутосинтетическая функция при размножении клеток) и от ДНК через иРНК к белку (гетеросинтетическая функция при биосинтезе белка).
| Транскрипция |
| Трансляция иРНК Белок |
ДНК
Репликация
Такой путь передачи информации от ДНК, которая способна к самовоспроизведению, к иРНК и белку Ф. Крик (1958) назвал «центральной догмой молекулярной биологии».
Долгое время считалось, что передача генетической информации в обратном направлении, т.е. от РНК до ДНК, невозможна. Однако в 1970 г Г. Тимин сообщил, что у РНК-содержащих вирусов с помощью ферментов ревертаз может синтезироваться на РНК комплементарная цепь ДНК для внедрения в геном клетки-хозяина. Спустя пять лет (1975 г) Р. Дульбеко, Г. Тимин и Д. Балтимор показали, что процессы обратной транскрипции могут происходить в клетках разных организмов от бактерий до млекопитающих и человека. Было установлено, что на ранних стадиях эмбриогенеза в клетках амфибий происходит резкое увеличение числа копий генов рибосомальной РНК (амплификация генов) с помощью обратной транскрипции.
Наличие ревертаз в нормальных клетках разных по систематическому положению организмов свидетельствуют о широком распространении процессов обратной транскрипции в живой природе и необходимости дополнения центральной догмы биологии этим важным обстоятельством.
| транскрипция |
| трансляция |
| Обратная транскрипция |
ДНК иРНК белок
РЕПЛИКАЦИЯ
Из приведенной схемы видно, что генетическая информация заключенная в нуклеиновой кислоте может передаваться лишь в направлении нуклеиновая кислота → белок. Передача информации в обратном направлении – от белка к нуклеиновой кислоте – невозможна.
В потоке биологической информации участвует ДНК хромосом, молекулы иРНК, переносящие информацию в цитоплазму, цитоплазматический аппарат трансляции (рибосомы и полисомы, тРНК, ферменты).
На завершающем этапе потока информации в клетке полипептиды, синтезированные на рибосомах, преобретают вторичную, третичную, четвертичную структуру и используются в качестве ферментов, строительных блоков, антител и др. (рис 9.1.)

Рис. 9.1. Поток биологической информации в клетке
Благодаря потоку информации клетка приобретает характерную для нее морфофункциональную организацию, поддерживает ее во времени и передает ряду поколений.
Передача качественно полноценной информации в ряду поколений обеспечивается путем воспроизведения (репликации идентичных двойных спиралей ДНК), а использование этой информации для организации клеточных функций путем биосинтеза белка в результате экспрессии генов.
9.2 Основные этапы экспрессии генов
(реализации генетической информации)
Экспрессия генов – совокупность биохимических процессов, в результате которых проходит преобразование генетической информации, представленной в последовательности нуклеотидов ДНК, в структуру молекулы иРНК, а затем в последовательность аминокислот в молекуле белка. Реакции синтеза иРНК и белка, осуществляются по матрицам (ДНК и РНК соответственно). Поэтому они получили название реакций матричного синтеза.
| Трансляция |
Транскрипция

9.2.1. Транскрипция – процесс перенесения (переписывания) генетической информации с ДНК на РНК. Матрицей для синтеза РНК служит только одна из двух цепей ДНК, так называемая кодогенная или матричная (она же смысловая) «+» цепь ДНК. Транскрипция происходит не на всей молекуле ДНК, а лишь на участке, отвечающем определенному гену. В процессе транскрипции образуются РНК всех трех типов – информационная (матричная), рибосомальная, транспортная.
Транскрипция состоит из множества молекулярных процессов, которые условно разделяют на этапы: инициация, элонгация, терминация.
Инициация транскрипции – начало синтеза РНК (запуск транскрипции), регулируется специфическими белками, активность которых зависит от метаболических процессов и нужд клетки.
Инициирует процесс транскрипции фермент РНК-полимераза, которая связывается с промоторным участком ДНК, а после это обеспечивает синтез РНК.
Промоторы прокариот и эукариот содержат короткие универсальные последовательности нуклеотидов, которые распознаются РНК-полимеразами и служат местом их присоединения. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Фермент геликаза разрывает водородные связи между азотистыми основаними параллельных цепей ДНК и они в этом месте расходятся. На одной из них (кодогенной) РНК-полимераза осуществляет синтез РНК по принципу элонгации (рис. 9.2.).

Рис.9.2. Схема синтеза мРНК
Матрицей для транскрипции мРНК служит кодогенная цепь ДНК, обращенная к ферменту своим 3ꞌ-концом
Элонгация – последовательное присоединение свободных нуклеотидов к кодогенной цепи ДНК по принципу комплементарности (А-У, Г-Ц) и соединение их в единую цепь при помощи РНК-полимеразы в полирибонуклеиновою цепочку.
Процесс элонгации требует присутствия ионов Mg2+ или Mn2+. Транскрипцию катализируют три разных типа полимераз. Это большие ферменты с четвертичной структурой. Первый тип синтезирует большие рибосомные РНК (рРНК), второй – транскрибирует гены, на которых закодированы полипептиды (образуется иРНК); третий - синтезирует тРНК и малую рРНК.
Терминация – завершение синтеза РНК в участке-терминаторе, который узнаётся РНК-полимеразой при участии особых белков факторов-терминации. В этом участке РНК-полимераза отделяется от матрицы ДНК от вновь синтезированной РНК. Фрагмент молекулы ДНК, который находится между промотором и терминатором образует единую транскрипцин - транскриптон.
В результате процесса транскрипции синтезируются разные виды РНК. Эти молекулы для своей функциональной активности в большинстве случаев проходят этап тех или иных модификационных изменений. Этап «созревания РНК», в процессе которого первичный РНК-транскрип (про-иРНК) превращается в молекулу функционально активной зрелой иРНК, получил название процессинга.
9.2.3. Процессинг как промежуточный этап экспрессии гена у эукариот
Напомним, что ген эукариот состоит из некодирующих участков интронов и кодирующих последовательностей нуклеотидов экзонов. При экспрессии генов, кодирующих структуру белка в результате транскрипции, которая заканчивается в зоне терминации, образуется про-иРНК (РНК-предшественница информационной); синонимы: незрелая РНК, гетерогенная РНК (гя-РНК). Она копирует всю нуклеотидную последовательность гена от промотора до терминатора, включая экзоны и интроны. Новообразованная про-иРНК подвергается тут же (в ядре) процессингу.
Посттранскрипционный процессинг – совокупность реакций «созревания» РНК, в результате которых первичная РНК-транскрипт (про-иРНК) превращается в зрелую функционально активную иРНК. Среди этих реакций выделяют сплайсинг, метилирование азотистых основ, кэпирование, полиаденилирование.
Сплайсинг – вырезание интронов ферментами – рестриктазами и соединение экзонов ферментами лигазами.
Метилирование азотистых оснований иРНК – присоединение до азотистых оснований метильных групп обеспечивает устойчивость иРНК до ферментов, которые могут ёё разрушить.
Кэпирование – процесс формирования «колпачка» или кэп-сайта за счет присоединения до 5/-конца иРНК молекулы ГТФ (гуанозинтрифосфата). Образование кэп-сайта («колпачка») обеспечивает узнавание молекул иРНК малыми субъеденицами рибосом.
Полиаденилирование – процесс присоединения к 3/ концу иРНК последовательности из 100-250 остатков адениловой кислоты, (поли-А), что способствует перемещению зрелой иРНК с ядра в цитоплазму. иРНК, которая образуется в результате процессинга схематически можно представить так:
Структура иРНК 1 – «кэп»; 2 – поли-А-участок; 3 – копии экзонов
Процессы модификации тРНК и рРНК происходят и у прокариот, что позволяет рассматривать их как прокариотический процессинг.
Обычно про-РНК в несколько раз (иногда в десятки) больше количества иРНК. Если про-РНК составляет около 10% генома, то иРНК только 1-2% генома.
У прокариот иРНК, которая образуется при транскрипции, процесинга не претерпевает.
Процессинг ядерных рРНК и тРНК проходит по другим схемам без просессов кэпирования и полиаденилирования, хотя сплайсинг часто происходит. Процессы модификации тРНК и рРНК происходят у прокариот, что позволяет рассматривать их как прокариотический процессинг.
Молекулы иРНК (после процессинга у эукариот и без процессинга у прокариот) участвуют в другом матричном процессе – трансляции (биосинтезе белка), который проходит на рибосомах.
9.2.4. Функциональная рибосома состоит из большой и малой субъединицы и молекулы иРНК. Рибосома имеет два активных участка – А (аминоацильный) и П (пептидальный), которые показаны на рис. 9.3.
| П |
| А |
| Малая субъединица |
| Большая субъединица |
| Рис. 9.3. Структура рибосомы А – аминоацильный участок П – пептидальный участок |
В пептидальном участке аминокислоты связываются пептидной связью и происходит наращивание полипептида, а к аминоацильному участку прикрепляются тРНК с активированными аминокислотами.
Основная функция рибосом – сборка белковых молекул из аминокислот, доставляемых к ним транспортными РНК (тРНК)
9.3 Трансляция
Трансляция – это процесс синтеза полипептидной цепочки на нити иРНК.
При трансляции происходит декодирование иРНК, в результате чего информация с языка последовательности оснований иРНК переходит на язык аминокислотной последовательности белка.
В процессе трансляции выделяют следующие стадии:
Стадия активации аминокислот. Активация свободных аминокислот с помощью особых ферментов аминоацил-тРНК-синтаз в присутствии АТФ. Активированная кислота присоединяется только к определённой тРНК с образованием комплекса аминоацил-тРНК (аа-тРНК). Процесс узнавания транспортной РНК своей аминокислоты называется рекогницией. Специфичность тРНК к аминокислоте определяется её антикодоном, а не аминокислотой. Молекула тРНК имеет сложную конфигурацию и похожа по форме на лист клевера. На верхушке расположен триплет свободных нуклеотидов (антикодон), которые по своему генетическому коду соответствуют определенной аминокислоте. На другом конце имеется участок для соединения с аминокислотой (рис. 9.4.)

Рис. 9.4..Схема биосинтеза белка
Инициация синтеза белка начинается с присоединения лидирующего 5/ конца иРНК до малой субъединице рибосомы. Соединение происходит так, что стартовый кодон на иРНК (всегда АУГ) располагается в области недостроенного П участка (рис. 9.5.). Далее инициирующая тРНК несущая метионин (аминоацил-тРНК с присоединенным метионином), связывается своим антикодоном со стартовым кодоном АУГ. Следующим шагом является объединение большой и малой субъединицы и «достраивание» П и А участков.
К концу фазы инициации в П участке расположен аминоацил-тРНК, связанная с метионином, а в А участке рибосомы располагается следующий за стартовым кодон иРНК.
Процессы инициации синтеза белка катализируются особыми белками – факторами инициации. По завершении образования инициирующего комплекса, состоющего из рибосомы, иРНК, инициирующей аминоацил – тРНК, эти факторы отделяются от рибосомы.

мРНК
Рис. 9.5. Инициация белкового синтеза:
1 – соединение малой субчастицы рибосомы с мРНК; тРНК, несущая метионин, присоединяется к стартовому кодону (АУГ) на мРНК и располагается в недостроенном П-участке; ІІ – соединение большой и малой субчастиц рибосомы с образованием П- и А- участков; следующий этап связан с размещением в А-участке аминоацил-тРНК, в соответствии с расположенным в нем кодоном мРНК, - начало элонгации; ак - аминокислота
Элонгация – это последовательное включение аминокислотных остатков в состав растущей полипептидной цепи (рис. 9.6.). Начинается из связывания антикодона очередной молекулы аминоацил-тРНК с кодоном, который расположен в свободном А-участке рибосомы. В результате на рибосоме оказывается две аминокислоты, между которыми образуется пептидная связь (0=С-N-H). Первая тРНК освобождается от аминокислоты и покидает рибосому. Рибосома перемещается вдоль нити иРНК в направлении 5/ - 3/ на один триплет, тем временем 2-я аа-тРНК, нагружается аминокислотами, перемещается в П участок, освобождая А-участок., который заполняет следующая 3-я аа-тРНК. Таким способом присоединяются 4-я, 5-я и т.д. аминокислоты, принесенные своими тРНК.
Таким образом, каждый акт элонгации состоит из трех этапов: 1) узнавание кодона; 2) образование пептидной связи; 3) транскрипции.
Все это повторяется многократно до тех пор, пока рибосома не дойдет до стоп кодона на РНК.
Терминация, т.е. окончание синтеза полипептида, происходит тогда, когда на рибосому попадает один из терминирующих («nonses») кодонов: УАА, УГА, УАГ. Стоп-кодоны не распознаются молекулами тРНК. Фактор терминации (специальный белок) присоединяется к этому кодону и блокирует дальнейшее передвижение рибосомы. Рибосома отсоединяется от иРНК и распадается на две субъединицы. Полипептид выходит в цитоплазму, где происходит его «созревание». При этом теряются некоторые концевые аминокислоты, формируется вторичная, третичная, или четвертичная структура.

Рис. 9.6. Фаза элонгации в синтезе белка
1-й этап аминоацил –тРНК присоединяется к кодону, расположенному в А-участке; 2-й этап – между аминокислотами, расположенными в А- и П- участка, образуется пептидная связь: тРНК, расположенная в П- участке, освобождается от своей аминокислоты и покидает рибосому; 3-й этап – рибосома перемещается по м РНК на один кодон так, что тРНК, нагруженная пептидной цепочкой, переходит из А-участка в П-участок; свободный А-участок может быть занят соответствующей аминоацил-тРНК
Синтез белковых молекул происходит непрерывно и идет с большой скоростью: синтез одной молекулы белка длится всего 3-4 сек. На одной молекуле иРНК может располагаться несколько рибосом (такое образование носит название полисома), что позволяет осуществлять синтез нескольких полипептидных цепей одновременно.
Об интенсивности процессов синтеза белков в организме человека свидетельствует следующее: половина белков тела человека (в среднем в нём около 17 кг белка) обновляется за 80 суток. За всю жизнь белки в организме человека обновляются около 200 раз.
9.5. Регуляция экспрессии генов
Клеточная ДНК несёт в себе генетическую программу, необходимую для синтеза сотен различных белков. Однако, в каждый данный момент клетка синтезирует только те белки, которые ей нужны в это время. Например, при выращивании кишечной палочки на питательной среде, не содержащей лактозы, её клетки содержат незначительное число (меньше пяти) молекул фермента лактазы, разлагающего лактозу на галактозу и глюкозу. При добавлении в питательную среду лактозы бактериальные клетки в течении 2-4 минут синтезируют больше 5 тыс. молекул лактозы. При удалении из среды лактозы синтез лактазы прекращается. Вещества, индуцирующие синтез ферментов, которые их разлагают называются индукторами (в данном примере индукторами является углевод лактоза).
Эти опыты свидетельствуют о том, что клетки бактерий способны регулировать свою генетическую активность для того, чтобы приспосабливаться к изменяющимся условием окружающей среды.
9.5.1. Регуляция экспрессии генов у прокариот
Регуляция генной активности у прокариот в основном осуществляется на уровне транскрипции. В 1961 г. французские ученые (будущие нобелевские лауреаты) Ф. Жакоб и Ж. Моно, разработали концепцию оперона для объяснения механизма «включения» или «выключения» тех или иных генов в зависимости от потребности клетки в определённых веществах, синтез которых контролируют эти гены. В дальнейшем эта концепция получила подтверждение в многочисленных экспериментах, показавших, что оперонная регуляция (т.е. регуляция на уровне транскрипции) является основным механизмом регуляции активности генов прокариот и бактериофагов.
Оперон - единица считывания генетической информации, транскрипция которой осуществляется на одну молекулу информационной РНК (иРНК) под контролем белка-репрессора. Основу оперона составляют расположенные в линейном порядком структурные гены и ген – оператор (рис. 9.7.).

Рис. 9.7. Схема регуляции транскрипции у прокариот
Структурные гены - кодируют белки - ферменты, осуществляющие последовательные этапы биохимических реакций одного метаболического пути (например, реакцию расщепления лактозы).
Ген-оператор - управляет функционированием структурных генов оперона, т.е. “включает” или “выключает” их. Если этот ген свободен, то транскрипция структурных генов происходит, если он связывается с белком-репрессором, то она прекращается.
В состав оперона, кроме структурных генов, гена - оператора входит промотр с инициатором и термінатором.
Промотор с инициатором - место первичного присоединения РНК- полимеразы - фермента, катализирующего реакцию ДНК-зависимого-синтеза иРНК.
Терминатор - последовательность ДНК, находящаяся на конце оперона и ответственная за прекращения транскрипции.
Ген-регулятор - обычно находится на некотором расстоянии от оперона, постоянно активен и на основе его информации синтезируется белок- репрессор.
Белок - репрессор при отсутствии в клетке индуктора соединяется с оператором и блокирует транскрипцию, так как РНК - полимераза не может двигаться вдоль оперона и транскрипция структурных генов не происходит (рис. 9.6). Если в клетку поступает индуктор то он связывает белок-репрессор, в результате чего репрессор не может присоединится к оператору. Свободный оператор “открывает путь” РНК-полимеразе и все гены оператора транскрибируются (рис 9.6.).
В результате транскрипции образуется иРНК (полицистронная), которая затем идет в рибосомы, где синтезируются ферменты разлагающие индуктор. Когда все молекулы индуктора будут разрушены, освобождается белок – репрессор, который снова связывает ген – оператор, в результате чего робота оперона прекращается. При поступлении индуктора она снова возобновляется. В описаном примера роль индуктора («побудителя» синтеза своего фермента) играют молекулы вещества, поступающие в клетку. Но в качестве индукторов могут выступать продукты распада белка, гормоны и другие метаболиты, которые могут связываться с белком-репрессором и тем самым влиять на транскрипцию структурных генов.
Для каждого оперона имеется свой специфический индуктор. Например, для лактозного оперона индуктором является лактоза, фруктозного – фруктоза, для гистидинового – гистидин. Основным преимуществом оперонной регуляции экспрессии генов у прокариот является синхронизация активности генов одного кластера и способность быстро переключать метаболизм с одного субстрата на другой.
9.6. Регуляция экспрессии генов у эукариот
В эукариотических клетках, также как и у прокариотических, образование определенных ферментов индуцируется присутствием их субстратов.
У эукариот опероны не обнаружены. Гены, контролируещие один метаболитическтий путь, у эукариот часто разбросаны по всему геному. Большинство (если не все иРНК) у эукариот моноцистронные, тогда как у прокариот иРНК полицистронная.
У эукариот, которые имеют нуклеосомную организацию хроматина, сложное строение гена, большое число факторов регуляции надклеточного уровня, включая факторы нервной и эндокринной систем, регуляция экспрессии генов очень сложная и осуществляется на многих этапах, ведущих от ДНК к белку.
9.6.1. Контроль на уровне транскрипции
Обобщённая схема регуляции транскрипции у эукариот разработана Г.П. Георгиевым (1972). В эукариотических клетках, также как и прокариотических, в основе регуляции лежат принцип обратной связи, но механизмы ее более сложные (рис.9.8.).
Единица транскрипции у эукариот называется транскриптоном, который состоит из неинформативной (акцепторной) и информативной (структурной) зон. Неинформативная зона включает промотор с инициатором и группу генов-операторов. Информативная зона представлена структурным геном, состоящим из экзонов и интронов (напомним, экзоны кодируют аминокислоты, а интроны - не кодируют аминокислот). Заканчивается транскриптон терминатором
Работу транскриптона регулирует несколько генов-регуляторов, несущих информацию для синтеза нескольких белков-репрессоров. Индукторами в клетках эукариот являются сложные вещества, для расщепления которых требуется несколько ферментов. Когда индукторы освобождают операторы от белков- репрессоров, РНК-полимезатора инициатирует транскрипцию на смысловой цепи структурного гена.

Рис.9.8. Схема регуляции транскрипции у эукариот
В результате транскрипция, которая заканчивается в зоне терминации, образуется про-информационная или гетерогенная ядерная РНК (про-иРНК, гя-РНК). Она контролирует всю нуклеотидную последовательность смысловой цепи ДНК структурного гена, т.е. включает экзоны и интроны. После этого гя-РНК претерпивает процессинг, или процес образования функционально активных и-РНК. Важным этапом процессинга является сплайсинг, в результате которого соеденяются экзоны и образуется иРНК (моноцистронная). Зрелая иРНК выходит их ядра в цитоплазму, соеденяется с рибосомами и служит матрицей для синтеза белка - фермента.
Кроме участка регуляторных генов, регуляция транскрипции у эукариот может осуществлятся путем спирализации – деспирализации хроматина. Хорошо известно, что деконденсация хроматина является необходимым условием для экспрессии гена.
На процессы «включения» и «выключения» генов оказывают влияния различные хромосомные перестройки, мигрирующие генетические элементы, изменяющие эффект положения гена.
Существенное влияние на транскрипцию генов у эукариот имеют гормоны (рис. 9.9.).
Контроль на уровне процессинга может осуществляться путем альтернативного сплайсинга. При этом «сшивание» отдельных информативных участков про-иРНК происходит по разному, в результате чего на основе одной и той же нуклеотидной последовательности одного гена образуются разные белки, состоящие из разных сочетаний одних и тех же аминокислот.
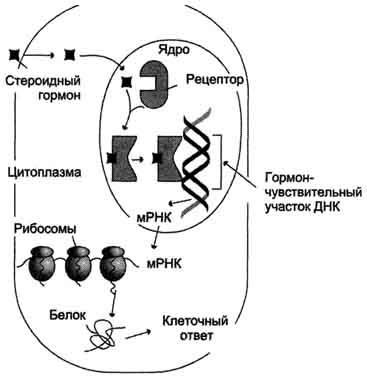
Рис. 9.9.
Действие стероидных гормонов, изменяющее транскрипцию. Гормон связывается с рецептором в клеточной мембране и вместе с ним направляется к клеточному ядру. Здесь он стимулирует транскрипцию определенных генов в мРНК.
Процессинг имеет избирательный характер, так как примерно половина гя- РНК распадается в ядре и не превращается в зрелую иРНК.
Контроль на уровне трансляции. В этом случае регуляция определяет, какие иРНК транслируются рибосомами и как часто они транслируются. На уровне трансляции обнаружена избирательная активность т-РНК, гормональные влияния на процессы синтеза белка.
Контроль на уровне посттрансляционной модификации белка. Процесс реализации генетической информации завершается посттрансляционной модификацией полипептида и превращение его в функционально активную молекулу белка. При этом происходит различные модификации аминокислот (ацетилирование, фосфорилирование), удаление некоторых из них, формирование вторичной, третичной, четвертичной структуры белка, функционально активных ферментов, гормонов и других веществ белковой природы. Все эти процессы происходят при участии большого числа факторов регуляции клеточного метаболизма.
Регуляция генной активности тесно связана с метаболизмом клетки, а также влиянием на нее целого ряда эндо- и экзогенных факторов и играет важную роль в сохранении динамического относительного постоянства состава и свойств клетки (гомеостаза). Ибо в связи с изберательной генной активностью сиснтезируются необходимые для жизнедеятельности клетки структурные белки и белки-ферменты через посредство которых осуществляются все прпоцессы ассимиляции и диссимиляции клетки.
9.7. Механизмы регуляции гомеостаза клетки
Гомеостаз клетки выражается в относительном постоянстве ее химического состава, осмотического давления, pH цитоплазмы, структурно-функциональной организации.
Включение и выключение процессов, обеспечивающих поддержание нормального химического состава и структурно-функциональной организации клетки, происходит в ней по принципу обратной связи автоматически. Такую регуляцию называют саморегуляцией, или авторегуляцией.
В основе авторегуляции деятельности клетки лежат процессы информации, т.е. сигналы. Сигналом служат изменение, возникшее в каком-либо звене клетки. В ответ на изменение включаются процессы, в результате которых это изменение устраняется.
Наглядным примером такой саморегуляции деятельности клетки может быть рассмотренный ранее механизм регуляции активности генов на уровне оперона.
Важное значение для поддержания гомеостаза клетки имеет изберательная проницаемость клеточной мембраны, процессы репликации и репарации ДНК, а также диплоидность эукариотических клеток.
Плазматическая мембрана позволяет проникать в клетки и выходить из них лишь определенным молекулам, и скорость обмена ими через мембрану строго регулируется возможностями диффузии, осмотическими и электрическими градиентами, активными механизмами, включающими транспортные системы мембран, и перемещениями мембранных структур, как, например, калий-натриевый насос.
Самое важное свойство живого – самовоспроизведение, в основе которого лежит процесс редупликации ДНК. Сам механизм этого процесса, при котором новая нить ДНК стоится строго комплементарно возле каждой из двух старых нитей, есть оптимальным для точной передачи информации. Точность этого процесса достаточно велика, но все-таки, хотя и редко, возникают ошибки при редупликации. В этом случае включается известный нам механизм самокоррекции, который устраняет возникшие нарушения в структуре ДНК.
Нарушение структуры молекулы ДНК может происходить и в ее первичных звеньях вне процесса репликации под воздействием эндогенных и экзогенных химических веществ, под влиянием физических факторов. В большинстве случаев происходит исправление повреждений ДНК с помощью системы репарационных ферментов. Репарация играет важную роль в восстановлении структуры генетического материала и сохранении нормальной жизнедеятельности клетки и ее гомеостаза. При нарушении механизмов репарации происходит нарушение гомеостаза, как на клеточном, так и на организменном уровнях.
Одним из механизмов сохранения гомеостаз является диплоидный набор хромосом в соматических клетках у эукариот. Большинство мутаций, которые часто имеют негативное влияние, есть рецессивными. Наличие доминантного аллеля обеспечивает полное или частичное угнетение в фенотипе рецессивной мутации.
Стабилизация сложной системы генотипа обеспечивается и явлениями полимерии, а также другими видами взаимодействия генов. У прокариот, которые имеют более примитивную организацию генотипа, наблюдается меньшая автономность их клеток от колебания внешней среды и более низкая стабильность самого генетического аппарата.
ЖИЗНЕННЫЙ ЦИКЛ И ДЕЛЕНИЕ КЛЕТКИ
10.1. Закономерности существования клетки во времени. Клеточный цикл.
Каждая клетка должна осуществлять все процессы от которых зависит её жизнь, т.е. поглощать питательные вещества, использовать их как пластический и энергетический материал, избавляться от конечных продуктов жизнедеятельности и, наконец, воспроизводить самую себя. В результате этих процессов клетка постоянно изменяется, осуществляется её онтогенез, который называется жизненным циклом клетки, или клеточным циклом.
Клеточный цикл – период существования клетки от момента её образования путём деления материнской клетки до собственного деления или смерти. У одноклеточных организмов клеточный цикл совпадает с жизнью особи. В многоклеточном организме клетки специализированы, т.е. имеют строго определённое строение и функции. В зависимости от специализации они имеют разную продолжительность жизни. Например, высокоспециализированные нервные и мышечные клетки после завершения эмбрионального периода развития перестают делится и функционируют на протяжении всей жизни организма. Другие клетки (костного мозга, эпителия тонкого кишечника, эпидермиса) в процессе выполнения своей специфичной функции быстро погибают и на смену им приходят новые, образовании путём деления клетки.
Все клетки возникают в результате деления уже существующих. Вновь образование клетки приобретают способность к делению после некоторого периода роста, удвоения ДНК, клеточных структур (митохондрий, хлоропластов, центриолей и др.).
Совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и в период деления называется митотическим циклом (от названия основного типа деления - митоза).
В непрерывно размножающихся клетках (например, эмбриональных) клеточный цикл совпадает с митотическим и длится от 10 до 50 ч. В других, частично специализированых клетках, митотический цикл составляет лишь часть их жизненного цикла. В жизненный цикл этих клеток включается период выполнения клеткой специфических функций, а также периоды покоя (рис.10.1).

Рис. 10.1. Жизненный цикл клетки многоклеточного организма.
I — митотический цикл; II — переход клетки в дифференцированное состояние; III— гибель клетки:
G1 — пресинтетический период, G2 — постсинтетический (предмитотический) период, М —митоз, S — синтетический период, R1 и R2 — периоды покоя клеточного цикла; 2с —количество ДНК в диплоидном наборе хромосом, 4с —удвоенное количество ДНК
Из Рис. 10.1. видно, что в периоде покоя судьба клетки не определена: она может начать подготовку к митозу, или же приступить к специализации в определенном функциональном направлении.

10.2 Изменение клетки в митотическом цикле
Митотический цикл - это время существования клетки от деления до деления. Состоит из интерфазы и митоза (рис.10.2.)
Интерфаза - это период между двумя последовательными делениями клетки. По продолжительности она занимает большую часть митотического цикла. В интерфазе происходят синтетические процессы, связанные с подготовкой клетки к делению.
В клетках, утративших способность к делению (напр. нейронах, эритроцитах), интерфаза – это период от последнего митоза и до смерти клетки.
Интерфаза включает три последовательных периода: пресинтетический (или постмитотический), синтетический, постсинтетический (премитотический).
Пресинтетический период (G1) - наиболее длительный период митотического цикла. В этот период клетка растет, выполняет свои функции; в ней усиленно образуются РНК и белки, АТФ. Каждая хромосома состоит из одной хроматиды. В состав хроматиды входят молекула ДНК, гистоновые и негистоновые белки. Содержание генетического материала в клетке соответствует 2n2c; n – гаплоидный набор хромосом, с – содержание ДНК в гаплоидном наборе.
Синтетический период (S) - период синтеза ДНК и репликации хромосом. В этот период происходит удвоение хроматид и содержание генетического материала в клетке становится 2n4c. В S-период продолжается синтез РНК, белков, АТФ.
Постсинтетический период (G2) - период формирования структур, необходимых для процесса деления клетки. Продолжается интенсивный синтез РНК и белков, запасается энергия в виде АТФ. К концу этого периода удваиваются центриоли. Содержание ДНК остается прежним (2n4c).
После того как в клетке завершаются биохимические процессы подготовки к делению, начинается заключительный этап митотического цикла – митоз.
Митоз – основной способ деления эукариотической клетки. В нём выделяют 4 следующие друг за другом фазы: профазу, метафазу, анафазу, телофазу (рис. 10.3.).
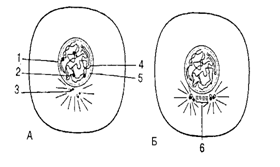
Интерфаза. Хромосомы распределены по всему ядру в виде рыхлой массы. Ядро и ядрышко отчётливо видно.

Профаза. Идёт процесс спирализации хромосом, исчезают ядрышки, разрушается ядерная оболочка. Центриоли расходятся к полюсам. Образуется веретено деления из микротрубочек. Содержание генетического материала - 2n4c.



Рис. 10.3. Схема митоза. А,Б – интерфаза; В,Г – профаза; Д – метафаза; Е – анафаза; Ж,З – телофаза:
1 – центромера; 2 – ядрышко; 3 – центриоль; 4 – хромосома; 5 – ядерная оболочка; 6 - веретено
Метафаза. Хромосомы достигают максимальной спирализации и выстраиваются в экваториальной плоскости. Содержание генетического материала - 2n4c.
Анафаза. Центромера делится пополам и хроматиды начинают синхронно расходится к полюсам клетки. С этого момента они становятся самостоятельными дочерними хромосомами. Содержание генетического материала - 4n4c.
Телофаза. Происходит деспирализация хромосом. Вокруг хромосом у каждого полюса формируется ядерная оболочка, в ядрах образуются ядрышка. Разрушается веретено деления. Одновременно идёт деление цитоплазмы. Дочерние клетки имеют диплоидный набор однохроматидных хромосом - 2n2c
10.2.1 Биологическое значение митоза определяется образованием новых клеток с идентичным содержанием генетической информации. Благодаря этому сохраняется преемственность в многочисленных поколениях клеток, поддерживается постоянство кариотипов особей вида.
Воспроизведение клеток приводит к увеличению их числа, обеспечивая этим процессы роста, развития, регенерации, замещения отмирающих клеток (эпителия кишечника, эпидермиса кожи, семенников и кроветворных органов).
В результате деления соматических клеток формируется, сохраняются и воспроизводятся клеточные популяции.
Клеточная популяция – это совокупность однородных клеток определённого вида ткани, которая поддерживается на относительно постоянном уровне, необходимом для выполнения тканью присущей ей функции. У многих таких популяциях жизненный цикл клеток ограничен небольшим сроком, поэтому клеточные популяции поддерживаются благодаря митозу.
Многие виды одноклеточных растений и животных размножаются бесполым путём при помощи одного лишь митотического деления клеток.
10.2.2. Митотическая активность ткани – это число клеток, которые делятся митозом на 1000 изученных клеток на гистологическом препарате.
У разных организмов и в разных тканях митотическое деление клеток протекает с различной скоростью - с наибольшим у бактерий и у зародышей многоклеточных организмов и с наименьшей - у высоко дифференцированных тканях.
Существует суточный ритм митотической активности. У животных, которые ведут ночной образ жизни, максимум митозов отмечается в утренние часы, а минимум - в ночные. Иная картина наблюдается у многих животных с дневной активностью: у них более высокая митотическая активность в большинстве органов наблюдается в ночное время суток, чем в дневное.
Факторами, от которых зависит пролиферация (деление) клеток, являются биологически активные вещества, среди которых особое место занимают гормоны, продукты разрушения тканей.
Так адреналин препятствует вступление клетки в митоз, тироидин в небольших дозах стимулирует пролиферацию (деление) клеток многих тканей, однако в больших дозах – оказывает обратный эффект.
Половые гормоны – эстрогены стимулируют пролиферацию клеток слизистой оболочки матки после менструации. Стимулирующие влияние на митозы имеют продукты разрушения тканей, что имеет значение для осуществления процессов регенерации, восстановления целостности тканей и органов многоклеточного организма.
Митоз чувствительный к действию различных физических и химических факторов, зависит от режима питания организма, его функционального состояния и благополучия.
Рентгеновские, или гамма – лучи, некоторые химические вещества угнетают митоз. Данный эффект угнетения митоза используют для прекращения или замедления пролиферации клеток раковых опухолей.
Все новые клетки, возникшие в результате деления уже существующих надвое, становятся способными к митозу лишь после некоторого периода роста.
Рост клетки – это результат преобладания в ней процессов ассимиляции (синтеза, накопления веществ) над процессом диссимиляции (трата, использование веществ). В процессе роста происходит изменение соотношения объёмов ядра и цитоплазмы.
При определённых значениях этого показателя рост клетки становится невозможным без дополнительных приспособительных изменений, которые компенсируют неблагоприятное для обмена веществ соотношение ядра и цитоплазмы.
Приспособительными изменениями могут быть:
· полиплоидизация и увеличение объёма ядра (макронуклеус у инфузорий, ядра у позвоночных и человека);
· увеличение количества ядер (многоядерные формы простейших, волокна скелетных мышц).
В большинстве случаев клетки после достижения определённого специфического для данного вида ткани – размера делятся митотически. Вновь образованные клетки могут снова расти до исходных размеров материнской клетки.
Таким образом, поочередные митозы являются предпосылкой для роста клеток, а рост клеток в большинстве случаев обуславливает их деление.
Факторами роста могут быть белки, полипептиды, стероиды.
Полипептидные факторы роста выделены в суперсемейства:
1. суперсемейство инсулиноподобных факторов роста (инсулин, проинсулин, релаксин и др.);
2. суперсемейство эпидермальных факторов роста;
3. суперсемейство бомбензину (бомбензин, литорин, литорян, нейротензин);
4. суперсемейство факторов роста фибробластов (ФРТ);
5. суперсемейство трансформирования факторов роста;
6. суперсемейство факторов роста тромбоцитов (ФРТ);
7. цитокинины.
Обычно под фактором роста понимают полипептиды, которые способные связываться со специфическими рецепторами клеток-мишеней и оказывать на них определенное регулирующее действие. В соответствии с этими критериями до факторов роста клетки относят интерфероны, интерлейкины, факторы некроза, опухолей и некоторые другие виды полипептидов, принимающих участие в регуляции клеточной пролиферации.
10.2.3 Нарушение митоза. Эндомитоз. Политения
Различные внешние и внутренние факторы (радиация, вирусы, алкоголь, наркотики, нервные стрессы, некоторые лекарства) могут вызывать нарушения в течении митоза, что ведёт к появлению несбалансированных наборов хромосом и разным патологиям клеток. С патологическими митозами связано возникновение многих болезней человека, особенно если они возникают на ранних стадиях эмбрионального развития. Считают, что нарушения митоза - одна с причин злокачественного перерождения клеток.
На основе митоза возникли механизмы, с помощью которых в том или ином органе количество наследственного материала может быть увеличено при сохранении постоянства числа хромосом. Примером таких механизмов может быть эндомитоз и политения.
Эндомитоз. При эндомитозе после удвоения ДНК и репродукции хромосом, деление ядра и цитоплазмы не происходит. Иными словами, эндомитоз – это процесс воспроизведения хромосом без формирования веретена деления клетки при сохранении ядерной оболочки.
Эндомитоз приводит к кратному увеличению числа хромосом по сравнению с диплоидным, т.е. сопровождается полиплоидией. Эндомитоз чаще всего встречается в клетках тканей, которые интенсивно функционируют (например, клетках печени, мышечных волокнах).
Политения заключается в кратном увеличении содержания ДНК в хромосомах при сохранении их диплоидного количества. Хромосомы приобретают большие размеры. Клетки с политенными хромосомами имеют увеличенное количество наследственного материала, что повышают их функциональные возможности. Это явление наблюдается в клетках двукрылых насекомых, инфузорий, зародышевых мешков некоторых растений.
10.3 Жизнь клетки вне организма. Клонирование клеток
Соматические клетки эукариот, в том числе и человека, могут расти и размножатся в лабораторных условиях (in vitro) на специальных питательных средах, т.е. существовать в виде так называемых клеточных культур.
Благодаря быстрому размножению клеток на питательных средах можно в течении короткого времени получать их в необходимом количестве для цитогенетических, биохимических, иммунологических исследований.
В условиях in vitro изолированные эукариотические клетки чаще всего проходят ограниченное количество делений, а затем гибнут. Исключение составляют раковые клетки, которые способны расти неограниченно долго. Способность этих клеток к длительному размножению в культуре, несомненно, связана с неограниченной пролиферативной способностью раковых клеток в живом организме.
Размножение клеток в культуре происходит по механизму митоза и приводит к образованию клеточных клонов.
Клеточный клон – совокупность генетически однородных клеток (чистая линия клеток), являющихся потомками одной родоначальной клетки.
Клонирование клеток – получение чистой линии клеток в лабораторных условиях, даёт возможность проводить в генетически идентичных клетках биохимический анализ наследственно обусловленных процессов.
Хромосомный набор длительно поддерживающихся культур клеток отличается от набора нормальных клеток. При культивировании часто происходят потери хромосом, некоторые сегменты хромосом могут делитироваться, дуплицироваться или перемещаться. Возникающие аномалии можно использовать для картирования генов и других генетических исследований.
Наиболее значительные успехи в генетических исследованиях человека связаны с гибридизацией соматических клеток.
Гибридизация соматических клеток основана на слиянии совместно культивируемых клеток разных типов, в результате чего образуются гибридные клетки, содержащие функциональные хромосомы обоих родительских видов. Гибридизация возможна между соматическими клетками разных людей, а так же между клетками человека и животных (например, мыши, крысы, обезьяны, морской свинки).
Гибридные клетки при размножении обычно «теряют» определённые хромосомы. Например, в гибридных клетках «человек- мышь» при их делении постепенно утрачиваются все хромосомы человека, а в клетках «человек - крыса» - все кроме одной, хромосомы крысы.
Гибридизация соматических клеток in vitro в сочетании с биохимическими и молекулярно-генетическими исследованиями открывает новые подходы и перспективы в изучении локализации генов в хромосомах, механизмов первичного действия и взаимодействия генов, клеточной дифференцировки и изменчивости.
Методы генетики соматических клеток, основанных на размножении этих клеток в искусственных условиях, позволяют проводить точную диагностику наследственных болезней в пренатальном периоде.
10.4 Амитоз как нетипичный способ деления клетки
Деления клетки бывают прямым (амитоз) и непрямым (митоз, мейоз). Большинство соматических клеток делятся путём митоза.
Амитоз - прямое деление клетки, при котором не образуется аппарат деления и хромосомы в световом микроскопе неразличимы. Ядро делится путём кольцевой перетяжки.
При амитозе часто наблюдается только деления ядра. В этом случае возникают двух – и многоядерные клетки; если же за делением ядра следует деление цитоплазмы, то распределение ДНК и клеточных элементов происходит произвольно, неравномерно.
Амитоз наблюдается в простейших, клетках растений и животных. У человека такой тип деления характерен для клеток печени, хрящей, роговицы глаза.
Амитоз наблюдается также в некоторых высокоспециализированных клетках с ослабленной физиологической активностью, дегенерирующих клетках, либо при различных патологических процессах, таких как злокачественный рост, воспаление и т.п.
Клетки, образованные в результате амитоза, имеют измененный кариотип и чаще всего быстро гибнут (например, клетки зародышевых оболочек млекопитающихся).
Амитоз в отличие от митоза является более экономным способом деления, так как энергетические затраты при амитозе весьма незначительны.
10.5 Мейоз. Сходство и различия между митозом и мейозом
Мейоз – это такой способ клеточного деления, в результате которого из диплоидных (2n) соматических клеток половых желёз образуются гаплоидные гаметы (n).
В ходе дальнейшего оплодотворения ядра гамет сливаются и восстанавливается диплоидный набор хромосом. Таким образом, основное биологическое значение мейоза заключается в обеспечении постоянства числа хромосом в ряду поколений организмов данного вида при половом размножении. Важным следствием мейоза являются обеспечение генетического разнообразия гамет в результате независимого расхождения каждой пары гомологических хромосом и кроссинговера.
Мейоз включает два последовательных деления. Эти деления называются: первое деление мейоза (мейоз І) и второе деление мейоза (мейоз ІІ). В каждом из этих делений различают четыре стадии: профазу, метафазу, анафазу и телофазу.
Первое мейотическое деление называется редукционным, так как оно приводит к образованию из диплоидных (2n) клеток гаплоидных (n). При втором мейотическом делении гаплоидность клеток сохраняется. Поэтому его называют эквационным.
10.5.1 Особенности первого (редукционного) мейотического деления
Интерфаза І. Предшествующая мейозу интерфаза полностью аналогична митотической интерфазе. Происходит синтез АТФ, белков, РНК, увеличение числа органел и клетка увеличивается в размерах. Удвоение ДНК и хроматид происходит в течении S – периода. Формула генетического материала клеток вступающих мейоз соответствует - 2n4с, где n – число хромосом, с – количество ДНК в хромосомном наборе.
Профаза І. Наиболее сложная и длительная стадия мейоза. Помимо процессов аналогичных процессам митоза (спирализация хромосом, разрушение ядерной оболочки, исчезновение ядрышка, образование веретена деления), ключевое значение имеет коньюгация гомологических хромосом – синапс. Соединенные пары гомологических хромосом называется бивалентами. Гомологичные хромосомы связывает особая структура, образованная из белков кариоплазмы – синаптонемальный комплекс (СК). В бивалентах гомологичные хромосомы могут обмениваться гомологичными участками.
Такой процесс называется кроссинговером. Профазу І обычно подразделяют на 5 подстадий: лептонему, зигонему, пахинему, диплонему и диакинез (рис. 10.4.).

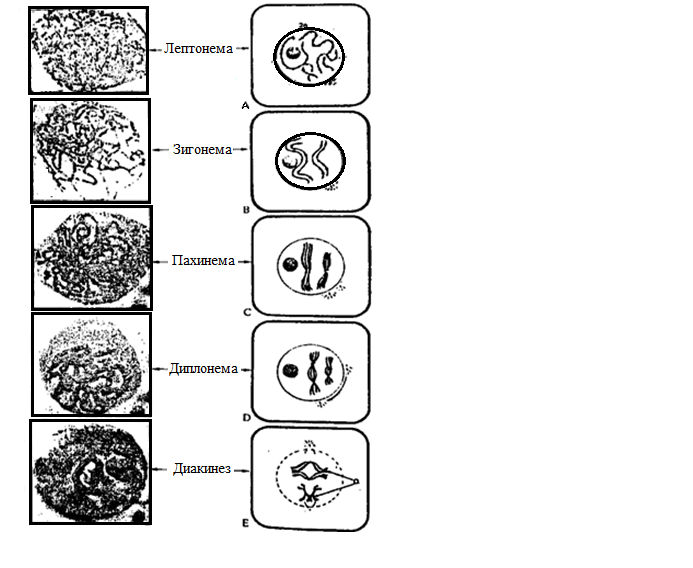
Рис. 10.4. Мейоз І (профаза І)
(формула генетического материала соответствует 2n4с)
Лептонема. Происходит спирализация хромосом, они укорачиваются и становятся видимыми как обособленные тонкие нити.
Зигонема. Начало (с отдельных участков) и завершение коньюгации гомологических хромосом. Хромосомы коньюгируют т.е. соединяются друг с другом, наподобие затяжки «молния». Такое соединение гомологических хромосом называется синапсом. Две сцепленные таким образом хромосомы называется бивалентом. Каждый бивалент состоит из 4 хроматид. Число бивалентов в клетке равно (у человека n=23).
Пахинема. Происходит укорочение и утолщение бивалентов (стадия толстых нитей) на фоне продолжающейся спирализации хромосом между гомологичными хромосомами происходит – кроссинговер – перекрест с обменом соответствующими участками. Участок, в которых две гомологические хромосомы обмениваются своим наследственным материалом при мейозе, называется хиазмой.
Диплонема. Гомологичные хромосомы начинают расходится (разрушается СК), но они связаны в областях прошедшего кроссинговера – хиазмах. Число хиазм бывает различным (обычно 2-3),в длинных хромосомах больше чем в коротких. Хиазмы показывают, что между хроматидами происходит кроссинговер.
Диакинез. Хромосомы достигают максимальной спирализации и интенсивно окрашиваются. Гомологичные хромосомы удерживаются вместе лишь в отдельных точках хиазм. Биваленты приобретают причудливую форму колец, крестов, восьмерок и т.д. Ядерная оболочка и ядрышко исчезают. Центриоли, если они есть, мигрируют к полюсам и затем образуют нити веретена.

Завершается формированием веретена деления клетки. Биваленты концентрируются в экваториальной плоскости клетки. Нити веретена деления прикрепляются до центромер гомологичных двуххроматидных хромосом, которые расположены по обе стороны от экваториальной плоскости.
Содержание генетического материала соответствует - 2n4c.

К полюсам клетки расходятся двуххроматидные хромосомы; при этом число хромосом у каждого полюса становится вдвое меньше, чем в материнской клетке (происходит редукция). Содержание генетического материала становится - n2c.

Обычно очень короткая. У полюсов клетки группируются гаплоидные наборы хромосом. Востанавливается структура ядра и ядерная оболочка. Происходит частичная деспирализация хромосом.
В конце телофазы І наступает цитокинез и образуется две клетки с гаплоидным набором двуххроматидных хромосом.
Формула генетического материала, образующихся дочерних клеток соответствует – n2c.
10.5.2. Особенности второго (эквационного) мейотического деления
Интерфаза ІІ (интеркинез). Интерфаза между мейозом І и мейозом ІІ обычно проходит быстро или отсутствует вообще. Её важное отличие от интерфазы, предшествующей мейозу І или митозу, состоит в том, что синтеза новой ДНК (т.е. репликации) в промежутке между первым и вторым мийотическими делениями не происходит. Вновь образованные дочерние клетки быстро вступают в мейоз ІІ, который происходит по механизму митоза.

Происходит незначительная спирализация двухроматидных хромосом, исчезает ядрышко и ядерная оболочка. Центриоли перемещаются к полюсам клетки, начинает формироваться веретено деления и перемещение хромосом до экваториальной плоскости. Содержание генетического материала
соответствует - n2c.

Хромосомы выстраиваются в экваториальной плоскости. Центромеры прикрепляются к микротрубочкам образованного веретена деления. Содержания генетического материала не изменяется - n2c.

Происходит разделение центромер и каждая хроматида становится самостоятельной хромосомой. Дочерние хромосомы отходят к противоположным полюсам клетки и содержание генетического материала у каждого полюса становится - nc.

Хромосомы деспирализуются, исчезает веретено деления, формируется ядрышко и ядерная оболочка, наступает цитокинез.

В телофазе образуется 4 гаплоидных клетки. Содержание генетического материала каждой клетки соответствует – nc.
Мейоз представляет единый процесс, что наглядно иллюстрирует рис.10.5.

Рис.10.5. Главные стадии мейоза
10.5.3. Сходство и различие между митозом и мейозом
Сходство процессов заключается в том, что биологическим смыслом каждого из них является деления генетического материала между дочерними клетками.
В митоз и мейоз вступают соматические клетки с одинаковым содержанием
генетического материала – 2 n4c.
В митозе и мейозе выделяют одинаковые фазы деления, которые характеризуются сходными процессами: спирализацией хромосом, преобразованием ядрышек, растворением ядерной оболочки, образованием ахроматинового веретена деления и др. Сходными являются детали строения хромосом и цитологические механизмы деления генетического материала.
Митоз – это такой способ клеточного деления, в результате которого из одной исходной клетки образуются две дочерние, в каждой из которых количество и объём генетической информации остаются неизменными, а масса ДНК уменьшается в два раза. Он происходит при делении соматических клеток и лежит в основе процессов роста, развития, восстановления, регенерации.
Мейоз – это такой способ клеточного деления, в результате которого из одной исходной клетки образуются 4 производных, каждая из которых содержит вдвое меньшее количество хромосом, в 4 раза меньшее количество ДНК и перекомбинированную генетическую информацию в сравнении с исходной клеткой. Он происходит в половых железах (зоне созревания) у животных и человека.
Мейоз является основным этапом формирования гамет, после слияния которых предотвращается удвоение числа хромосом в каждом новом поколении.
Важным следствием мейоза является обеспечение генетического разнообразия гамет в результате рекомбинации хромосом и кроссинговера.
Дата публикования: 2014-09-25; Прочитано: 9730 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
