 |
Главная Случайная страница Контакты | Мы поможем в написании вашей работы! | |
Обучение. Только простые узлы имеют настраиваемые веса
|
|
Только простые узлы имеют настраиваемые веса. Это веса связей, соединяющих узел с комплексными узлами в предыдущем слое и имеющих изменяемую силу синапсов, настраиваемую таким образом, чтобы выработать максимальную реакцию на определенные стимулирующие свойства. Некоторые из этих синапсов являются возбуждающими и стремятся увеличить выход узлов, в то время как другие являются тормозящими и уменьшают выход узла.
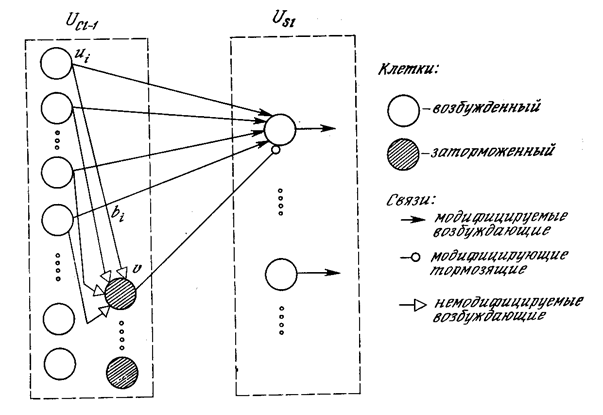
Рис. 10.11. Связи от сложных клеток одного уровня
к простым клеткам следующего уровня
На рис. 10.11 показана полная структура синаптических связей между простым узлом и комплексными узлами в предшествующем слое. Каждый простой узел реагирует только на набор комплексных узлов внутри своей рецептивной области.
Кроме того, существует тормозящий узел, реагирующий на те же самые комплексные узлы. Веса синапсов тормозящего узла не обучаются, – они выбираются таким образом, чтобы узел реагировал на среднюю величину выходов всех узлов, к которым он подключен. Единственный тормозящий синапс от тормозящего узла к простому узлу обучается, как и другие синапсы.
Обучение без учителя. Для обучения неокогнитрона на вход сети подается образ, который необходимо распознать, и веса синапсов настраиваются слой за слоем, начиная с набора простых узлов, ближайших ко входу. Величина синаптической связи от каждого комплексного узла к данному простому узлу увеличивается тогда и только тогда, когда удовлетворяются следующие два условия:
1) комплексный узел реагирует;
2) простой узел реагирует более сильно, чем любой из его соседних (внутри его области конкуренции).
Таким образом, простой узел обучается реагировать более сильно на образы, появляющиеся наиболее часто в его рецептивной области, что соответствует результатам исследований, полученных в экспериментах с котятами. Если распознаваемый образ отсутствует на входе, тормозящий узел предохраняет от случайного возбуждения.
Математическое описание процесса обучения и метод реализации латерального торможения аналогичны описанным для когнитрона, поэтому здесь они не повторяются. Необходимо отметить, что выходы простых и комплексных узлов являются аналоговыми, непрерывными и линейными и что алгоритм обучения предполагает их неотрицательность.
Когда выбирается простой узел, веса синапсов которого должны быть увеличены, он рассматривается как представитель всех узлов в плоскости, вызывая увеличение их синаптических связей на том же самом образе. Таким образом, все узлы в плоскости обучаются распознавать одни и те же свойства, и после обучения будут делать это независимо от позиции образа в поле комплексных узлов в предшествующем слое.
Эта система имеет ценную способность к самовосстановлению. Если данный узел выйдет из строя, будет найден другой узел, реагирующий более сильно, и этот узел будет обучен распознаванию входного образа, тем самым перекрывая действия своего отказавшего товарища.
Обучение с учителем. В работах [3] и [8] описано самоорганизующееся неуправляемое обучение. Наряду с этими впечатляющими результатами, были опубликованы отчеты о других экспериментах, использующих обучение с учителем [9]. Здесь требуемая реакция каждого слоя заранее определяется экспериментатором. Затем веса настраиваются с использованием обычных методов для выработки требуемой реакции. Например, входной слой настраивался для распознавания отрезков линий в различных ориентациях во многом аналогично первому слою обработки зрительной коры.
Последующие слои обучались реагировать на более сложные и абстрактные свойства до тех пор, пока в выходном слое требуемый образ не будет выделен. При обработке сети, превосходно распознающей рукописные арабские цифры, экспериментаторы отказались от достижения биологического правдоподобия, обращая внимание только на достижение максимальной точности результатов системы.
Реализация обучения. В обычных конфигурациях рецептивное поле каждого нейрона возрастает при переходе к следующему слою. Однако количество нейронов в слое будет уменьшаться при переходе от входных к выходным слоям. Наконец, выходной слой имеет только один нейрон в плоскости сложных узлов. Каждый такой нейрон представляет определенный входной образ, которому сеть была обучена.
В процессе классификации входной образ подается на вход неокогнитрона и вычисляются выходы слой за слоем, начиная с входного слоя. Так как только небольшая часть входного образа подается на вход каждого простого узла входного слоя, некоторые простые узлы регистрируют наличие характеристик, которым они обучены, и возбуждаются. В следующем слое выделяются более сложные характеристики как определенные комбинации выходов комплексных узлов. Слои за слоем свойства комбинируются во все возрастающем диапазоне; выделяются более общие характеристики и уменьшается позиционная чувствительность.
В идеальном случае только один нейрон выходного слоя должен возбудиться. В действительности обычно будет возбуждаться несколько нейронов с различной силой, и входной образ должен быть определен с учетом соотношения их выходов.
Если используется сила латерального торможения, возбуждаться будет только нейрон с максимальным выходом. Однако это часто является не лучшим вариантом. На практике простая функция от небольшой группы наиболее сильно возбужденных нейронов будет часто улучшать точность классификации.
Заключение
Как когнитрон, так и неокогнитрон производят большое впечатление с точки зрения точности, с которой они моделируют биологическую нервную систему. Тот факт, что эти системы показывают результаты, имитирующие некоторые аспекты способностей человека к обучению и познанию, наводит на мысль, что наше понимание функций мозга приближается к уровню, способному принести практическую пользу.
Неокогнитрон является сложной системой и требует существенных вычислительных ресурсов. По этим причинам кажется маловероятным, что такие системы реализуют оптимальное инженерное решение сегодняшних проблем распознавания образов. Однако с 1960 г. стоимость вычислений уменьшалась в два раза каждые два-три года, тенденция, которая, по всей вероятности, сохранится в течение как минимум ближайших десяти лет. Несмотря на то, что многие подходы, казавшиеся нереализуемыми несколько лет назад, являются общепринятыми сегодня и могут оказаться тривиальными через несколько лет, реализация моделей неокогнитрона на универсальных компьютерах является бесперспективной. Необходимо достигнуть тысячекратных улучшений стоимости и производительности компьютеров за счет специализации архитектуры и внедрения технологии СБИС, чтобы сделать неокогнитрон практической системой для решения сложных проблем распознавания образов, однако ни эта, ни какая-либо другая модель искусственных нейронных сетей не должны отвергаться только на основании их высоких вычислительных требований.
Литература
1. Blakemore С., Cooper G. F. 1970. Development of the brain depends on the visual enviroment. Nature 228(5270):477–78.
2. Fukushima К. 1975. Cognitron: A self–organizing mult ilayered neural network. Biological Cybernetics 20:121–36.
3. Fukushima К. 1980. Neocognitron: A self–organizing neural network model for a mechanism of pattern recognition uneffected by shift in position. Biological Cybernetics 36(4):193–202.
4. Fukushima К. 1981. Cognitron: A self–organizing multilayer neural network model. NHK Technical Monograph No. 30, pp. 1–25. Available from Nippon Hoso Kyokai (Japanese Broadcasting Corp.), Technical Research Labs, Tokio, Japan.
5. Fukushima K. 1984. A hierarchical neural network model for associative memory. Biological Cybernetics 50:105–113.
6. Fukushima К. 1986. A neural network model for selective attention in visual pattern recognition. Biological Cybernetics 55(1):5–15.
7. Fukushima К. 1987. A neural network for selective attention. In Proceedings of the IEEE First International Conference on Neural Networks, eds. M. Caudil and C. Butler, vol. 2, pp. 11–18. San Diego, CA:SOS Printing.
8. Fukushima K., Miyake S. 1982. Neocognitron: A new algorithm for pattern recognition tolerant of deformations and shifts in position. Pattern recognition 15(6): 455–69.
9. Fukushima К., Miyake S., Takayuki I. 1983. Neocog–nitron: A neural network model for a mechanism of visual pattern recognition. IEEE Transaction on Systems, Man and Cybernetics SMC–13(5):826–34.
10. Hubel D. H., Wiesel T. N. 1962. Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. Journal of Physiology 160:106–54.
11. Hubel D. H., Wiesel T. N. 1965. Reseptive fields and functional architecture in two nonstriate visual areas (18 and 19) of the cats. Journal of Neurophi–siology 28:229–89.
12. Hubel D. H., Wiesel. T. N. 1977. Functional architecture of macaque monkey visual cortex. Proceedings of the Royal Society, London. Ser. В 198, pp. 1–59.

Дата публикования: 2014-10-18; Прочитано: 632 | Нарушение авторского права страницы | Мы поможем в написании вашей работы!
